LOS CONIOPTERÍGIDOS DE LA PENÍNSULA IBÉRICA E ISLAS BALEARES (INSECTA: NEUROPTERIDA, NEUROPTERA: CONIOPTERYGIDAE)
Víctor J. Monserrat
Departamento de Zoología y Antropología Física. Facultad de Biología. José Antonio Nováis, 2, Universidad Complutense, 28040 Madrid (Spain). E-mail: artmad@bio.ucm.es
| |
RESUMEN
Se revisan las especies de coniopterígidos presentes en la fauna de la Península Ibérica e Islas Baleares donde, hasta la fecha, están representadas por 39 especies pertenecientes a 8 géneros. Partiendo de los datos generales conocidos, damos una diagnosis de la familia y citamos algunos elementos sobre su historial paleontológico y su distribución, y algunos datos sobre su morfología, su biología, sus estadios juveniles y comportamiento, y se comenta una breve introducción sobre el historial en el conocimiento de la familia, y en particular en la Península Ibérica y Baleares. Se propone revalidar a Coniopteryx (Holoconiopteryx) tullgreni Tjeder, 1930 sp. reval. como especie válida, se describe Helicoconis bachi n. sp. como nueva especie, y se mantiene, se discute y se argumenta nuestro criterio sobre la validez de Coniopteryx parthenia (Navás & Marcet, 1910) (sensu Meinander, 1972a) frente a Coniopteryx hoelzeli (sensu Günter, 1993). Se incluye una lista de las especies pertenecientes a la fauna de la Península Ibérica y Baleares, una clave de identificación de los imagos de las subfamilias, géneros y especies citadas, y una clave de sus larvas conocidas a nivel genérico. De cada una de estas especies se recopilan y se anotan todas las referencias bibliográficas existentes relativas a las especies de la zona estudiada, tanto con la denominación actualmente aceptada, como de sus sinonimias y/o cualquier otra combinación nomenclatural bajo las cuales hayan sido citadas. También de cada una de las especies se anotan los datos
conocidos y los ahora aportados (1.800 nuevos ejemplares de 32 especies) sobre su distribución general, aspectos de su morfología
externa y/o genital, así como de su variabilidad, estadios juveniles, biología, fenología y distribución altitudinal y geográfica
en la península y archipiélago balear, en base a un total de 12.215 ejemplares estudiados.
http://urn:lsid:zoobank.org:pub:2E2BB2E4-FF78-4B32-BC78-22427871D86C
Palabras clave: Insecta;
Neuroptera;
Coniopterygidae;
Revisión;
Taxonomía;
Faunística;
Biología;
Morfología;
Península Ibérica;
Baleares;
España;
Portugal.
|
| |
SUMMARY
The dusty-wings from the Iberian Peninsula and Balearic Islands (Insecta: Neuropterida, Neuroptera: Coniopterygidae)
The present contribution revises the dusty-wings species present in the fauna of the Iberian Peninsula and Balearic Islands
where, to date, is represented by 39 species belonging to 8 genera. Based on the known general data, a diagnosis of the family
and its paleontological history and distribution is given, and there are exposed some general data on its morphology and biology,
juvenile stages and behavior, and is commented a brief introduction on the history in the knowledge of the family, and in
particular on the Iberian Peninsula and Balearic Islands fauna. Coniopteryx (Holoconiopteryx) tullgreni Tjeder, 1930 sp. reval, is proposed as as a real valid species, Helicoconis bachi n.sp. is described as a new species, and our criterion on the validity of Coniopteryx parthenia (Navás & Marcet, 1910) (sensu Meinander, 1972a) as a real vallid species against Coniopteryx hoelzeli (sensu Günter, 1993) is reasoned, discussed and argued. All are included in a list of the species belonging to their fauna,
an imagoes identification key of the subfamilies, genera and species recorded in the Iberian Peninsula and Balearic Islands,
and a key, at genera level, of their larval stages. For every one of these species are compiled and annotated all the existing
bibliographic references, relating to the species present in the studied area, both with the currently accepted name, their
synonyms and/or any other nomenclatural combinations under which they have been recorded. Of each species is also detailed
the present general distribution is known, and now are given new recorded data (1.800 new specimens of 32 species), and its
external and/or genital morphology, as well as their variability, juvenile stages, biology, phenology and altitudinal and
geographic distribution in the Peninsula and Balearic Archipelago are compilled, on the basis of 12.215 studied specimens.
Key Words: Insecta;
Neuroptera;
Coniopterygidae;
Revision;
Taxonomy;
Faunistics;
Biology;
Morphology;
Iberian Peninsula;
Balearic Islands;
Spain;
Portugal.
|
IntroducciónTOP
GENERALIDADES: DIAGNOSIS, HISTORIAL PALEONTOLÓGICO Y DISTRIBUCIÓN
La familia Coniopterygidae representa una de las mejor conocidas desde el punto de vista de la Sistemática y Taxonomía, y
es, sin duda, una de las más uniforme y sorprendente dentro del orden de los neurópteros, tanto por la particular y “aberrante”
morfología de sus imagos en relación a otras familias (Figs. 1-9), con su pequeño tamaño, sus secreciones cerúleas que le dan este típico color blanquecino pulverulento, y su peculiar morfología
y venación, muy diferente a lo que habitualmente acontece en los neurópteros, como por la especialización de sus estadios
larvarios (Figs. 20-22), hechos que les permiten acceder a presas muy pequeñas y a nuevos recursos donde hallan una menor competencia.
Simbolizan otro ejemplo más de las sorprendentes y curiosas adaptaciones de los neurópteros a nichos específicos no ocupados
o menos explotados, que les ha permitido sobrevivir evitando la competencia y la presión ejercida por nuevos grupos de insectos
holometábolos más evolucionados, en este caso, reducir su tamaño y tener acceso a nuevos recursos, como hemos indicado, en
este caso a presas mucho más pequeñas.
Los coniopterígidos constituyen una familia de neurópteros bastante homogénea, particular e interesante, debido a su particular
morfología y anatomía que, en muchos aspectos, les separa del resto de los neurópteros, a su amplia distribución geográfica,
al elevado número de individuos que frecuentemente constituyen sus poblaciones, y especialmente por su utilización como aliados
de nuestros intereses, al tratarse de eficaces agentes de control de pequeños artrópodos fitófagos (Killington, 1936; New, 1975, 1986, 1989, 1991a, 1999, 2001a, 2001b; Nicoli Aldini, 2002; Viggiani, 1977; Dinkins et al., 1994; McEwen et al., 2001; Pantaleoni, 2007). Por ello, se trata de una de las familias de neurópteros que ha venido recibiendo mayor atención, aunque menor en comparación
con otras familias, de interés en el control biológico, sea Hemerobiidae o especialmente Chrysopidae, y por ello son ingentes
los datos y publicaciones existentes sobre la distribución, biología, fenología, morfología, histología, fisiología, estadios
juveniles y variabilidad de muchas de sus especies, sobre las que se han publicado varias monografías (Enderlein, 1906, 1908; Meinander, 1972a, 1990; McEwen et al., 2001; Sziraki, 2011), y existen cientos de artículos de interés faunístico o aplicado que citan estos insectos.
Desde los inicios de la Taxonomía y la Sistemática modernas, han existido numerosos investigadores que han tratado de ubicar
a los neurópteros (Neuropteroidea), en general, respecto a otros grupos de insectos, a sus tres órdenes reconocidos (Megaloptera,
Raphidioptera y Neuroptera) entre sí, y a los coniopterígidos en particular en relación a sus diferentes familias que iban
siendo segregadas y descritas (Brauer & Löw, 1857; Enderlein, 1905a, 1906, 1910a; Handlirsch, 1906, 1906-1908; Tillyard, 1916, 1926, 1932; Withycombe, 1925a; Hennig, 1969; Killington, 1936; Tjeder, 1957; Meinander, 1972a; Schlüter, 1986; Grimaldi & Engel, 2005, etc.), y que en general los consideran un grupo muy apartado del resto de los neurópteros, a veces considerados como “aberrantes”,
con ciertas vinculaciones con Ithonidae [familia de distribución australiana y neártica con escaso número de especies (4 géneros
y unas 15 especies) de larvas melolontiformes, ciegas y edáficas].
Muchos de los datos existentes en esta familia, tanto en la anatomía y morfología (mayoritariamente apomórficos, aunque también
plesiomorfos), bien de sus imagos (Figs. 1, 2, 10-14, 23, 24), como de sus estadios preimaginales (Figs. 5-9, 26-76) son muy particulares y únicos dentro del orden Neuroptera. En sus imagos (Figs. 1, 2, 10-14, 23, 24) es peculiar el tamaño reducido, cabeza hipognata, venación alar reducida, casi inexistente en el muy estrecho campo costal,
venas trasversales muy escasas, venación longitudinal escasamente bifurcada y nunca en sus extremos, ausencia de pterostigma
y trichosoros (pequeños espesamientos sobre el margen alar entre las bifurcaciones de las venillas), secreción de ceras merced
a glándulas hipodérmicas con las que se cubren casi por completo su cuerpo y sus alas, presencia de vesículas abdominales
evaginables (plicaturas) que recuerdan a las vesículas coxales o tubos ventrales evaginables de algunos apterigotas (p. ej.
Collembola, Microcoryphia, Zygentoma, Protura), presencia en los machos de distinta microestructura del espermatozoide, con
testículos solo funcionales en las fases larvarias con reservorio en los imagos y ausencia de espermatóforos pero con estructuras
genitales (pene) que inyectan el esperma desde el reservorio al interior de las vías genitales femeninas, ausencia de cercos,
y en las hembras tipo de ovariolas y vías genitales femeninas, ausencia de ovopositores, ausencia de callo cercal en la terminalia
de ambos sexos, etc., caracteres diferentes o que no existenten en el resto de neurópteros. En cuanto a sus estadios preimaginales
(Figs. 5-9, 26-76) destaca el labro bien desarrollado y proyectado hacia adelante, similar a lo acontecido en la primitiva familia Psychopsidae
(Withycombe, 1925a) y curiosamente seis y no ocho tubos de Malpigio (hecho que comparten con Megaloptera, Raphidioptera y Coleoptera) e histológicamente
muy diferentes al resto de neurópteros, mandíbulas y maxilas rectas, concentración a dos ganglios nerviosos abdominales, ausencia
de meconio (restos del contenido acumulado en el tubo digestivo que no ha podido ser eliminado por el ano con anterioridad,
debido a la falta de conexión entre el mesodeo y proctodeo, y elemento sólido que, como una única estructura, caracteriza
a todos los neurópteros tras su emergencia como imagos después de la fase pupal) debido a que esta familia, curiosamente,
no posee capacidad de producir membranas peritróficas, y este contenido se evacúa merced a varias gotitas de líquido viscoso,
etc. Esta combinación de caracteres, y al margen de muchos otros de naturaleza histológica (glándulas tegumentarias, gónadas,
gametos, tubos de Malpighi, mesodeo, etc.), son diferentes o no ocurren en los restantes neurópteros (Withycombe, 1923, 1925a; Killington, 1936; Zimmerman, 1957; Tjeder, 1957; Meinander, 1972a; Sziráki, 1996; Kubrakiewicz et al., 1998; Nelson et al., 2003; Zizzari et al., 2008, 2011; Grimaldi & Engel, 2005; Zimmermann et al., 2009; Winterton et al., 2010).
Por todo ello, esta familia ha sido de difícil y compleja vinculación filogenética en relación a las restantes familias de
neurópteros, incluso con otros grandes grupos de Neuropteroidea (Megaloptera) (Enderlein, 1910a y Karny, 1923, 1924 los incluyen en este orden, y Killington, 1936 menciona su aparente mayor relación con ellos que, salvo Ithonidae, con las restantes familias de neurópteros), y con frecuencia
e históricamente ha sido considerada como un grupo aislado e independiente del resto de los neurópteros (incluso a veces considerada
con categoría de superfamilia: Coniopterygoidea) (Krüger, 1923; Withycombe, 1925a; Carpenter & Lestage, 1928; Killington, 1936; Obenberger, 1958; Riek, 1970; Meinander, 1972a, 1979a; Popov, 1973; Henry, 1982; New, 1986, 1989; Schlüter, 1986; Dorokhova, 1987; Kubrakiewicz et al., 1998; Sziráki, 2007a; Tauber et al., 2009), próximos a Ithonidae (Crampton, 1921; Withycombe, 1925a; New, 1989), a Hemerobiidae-Chrysopidae (Handlirsch, 1906-1908; New, 2001a), a Osmylidae (Tillyard, 1919), a Sisyridae (H. Aspöck et al., 2001) o a Dilaridae (Zimmermann et al., 2009).
|
 Figs. 1-9.— Muestra de coniopterígidos, de arriba a abajo y de izquierda a derecha: Aleuropteryx, Coniopteryx, Semidalis, Conwentzia, 2-9: coniopterígidos en su ambiente natural, 2: imago de Semidalis sp., 3-4: puesta de Semidalis sp., 5-7: larvas de coniopterígidos devorando ejemplares y huevos de Saissetia oleae Olivier, 1791 (Insecta, Homoptera, Coccidae), 8: capullo de coniopterígido, 9: prepupa de coniopterígido (abierto artificialmente
el capullo) (foto 1, 5-9 de Manuel González Núñez, 2: de Isidro Martínez, 3, 4: de Víctor J. Monserrat). Figs. 1-9.— Muestra de coniopterígidos, de arriba a abajo y de izquierda a derecha: Aleuropteryx, Coniopteryx, Semidalis, Conwentzia, 2-9: coniopterígidos en su ambiente natural, 2: imago de Semidalis sp., 3-4: puesta de Semidalis sp., 5-7: larvas de coniopterígidos devorando ejemplares y huevos de Saissetia oleae Olivier, 1791 (Insecta, Homoptera, Coccidae), 8: capullo de coniopterígido, 9: prepupa de coniopterígido (abierto artificialmente
el capullo) (foto 1, 5-9 de Manuel González Núñez, 2: de Isidro Martínez, 3, 4: de Víctor J. Monserrat).
Figs. 1-9.— Dustywings sample, from top to bottom and to left to right: Aleuropteryx, Coniopteryx, Semidalis, Conwentzia, 2-9: habitus of dustywings in their natural environment, 2: imago of Semidalis sp., 3-4: eggs of Semidalis sp., 5-7: larvae of dustywings devouring specimens and eggs of Saissetia oleae Olivier, 1791 (Insecta, Homoptera, Coccidae), 8: dustywing cocoon, 9: prepupa of dustywing (cocoon artificially opened) (photograph
1, 5-9: by Manuel González Núñez, 2: by Isidro Martínez, 3, 4: by Víctor J. Monserrat).
|
|
Como está ocurriendo en la Zoología y la Entomología en general, con las nuevas herramientas moleculares que han venido apareciendo,
es muy probable que puedan modificar la situación de numerosas especies, y la posición de muchos taxa se va a ver, sin duda,
afectada en un futuro. Recientemente diversos estudios cladísticos y moleculares los han relacionado con Dilaridae y Sisyridae
dentro de los Hemerobiiformia, uno de los grandes y más extensos y diversos linajes que constituyen los Neuroptera s.str. (= Planipennia) Handlirsch, 1908 (Tauber & Adams, 1990; U. Aspöck, 1992, 1995, 2002, 2003; Aspöck & Aspöck, 1999, 2010, 2013; H. Aspöck et al., 2001; U. Aspöck et al., 2001, 2012; Haring & Aspöck, 2004; Zimmermann et al., 2009; Beutel et al., 2010, etc.), y que junto a los Raphidioptera Latreille, 1810 y los Megaloptera Latreille, 1810 forman el Superorden Neuropterida o Neuroptera s. l., que, en su conjunto, está constituido por unas 6.620 especies repartidas en estos tres órdenes citados y habitualmente reconocidos:
Megaloptera (c. 380 spp.), Raphidioptera (c. 241 spp.) y Planipennia o Neuroptera s. str. (c. 6.000 spp.) (H. Aspöck et al., 1980, 2001), cuyas relaciones aún son objeto de amplios estudios y debates (Ejs.: Tauber & Adams, 1990; Ren & Hong, 1994; Whiting et al., 1997; U. Aspöck et al., 2001, 2003, 2012, 2015; Winterton, 2003; Haring & Aspöck, 2004; Jepson & Penney, 2007; Zimmermann et al., 2009, 2011a, 2011b; Winterton et al., 2010; Haring et al., 2011; Peters et al., 2014, etc.). Los Hemerobiiformia estarían constituido por (Chrysopidae + Osmylidae + Polystoechotidae + Rapismatidae + Ithonidae),
grupo hermano de Hemerobiidae + [(Coniopterygidae + Sisyridae) + Dilaridae + (Mantispidae + Rhachiberothidae-Berothidae)]
(Ghilarov, 1962; Aspöck et al., 1980; Schlüter, 1986; U. Aspöck, 1992, 1995, 2002, 2003; Aspöck & Mansell, 1994; H. Aspöck et al., 2001; U. Aspöck et al., 2001, 2003, 2012; Haring & Aspöck, 2004; Engel & Grimaldi, 2007; Aspöck & Aspöck, 2008, 2010; Zimmermann et al., 2009, 2011a, 2011b; Tauber et al., 2009; Beutel et al., 2010, etc.), no sin falta de debates, de controversias y de consenso (Whalley, 1980; Tauber & Adams, 1990; Willmann, 1990, 1994a, 1994b; Ren & Hong, 1994; Whiting et al., 1997; Kubrakiewicz et al., 1998; Grimaldi, 2000; Winterton, 2003; Grimaldi & Engel, 2005; Zizzari et al., 2008, 2011; Zimmermann et al., 2009, 2011a, 2011b; Winterton et al., 2010; Haring et al., 2011; Peters et al., 2014; Morinière et al., 2014, etc.).
Más recientes estudios moleculares (Wang & Liu, 2007) han aportado nuevos datos que, desde nuestro punto de vista requieren mayor discusión, y otros estudios, en particular Zimmermann
et al. (2009) quienes los asocian a Dilaridae como grupo hermano frente a Sisyridae, y Winterton et al. (2010) quienes refutan la inclusión de Coniopterygidae dentro del clado [(Coniopterygidae + Sisyridae) + Dilaridae + (Mantispidae
+ Rhachiberothidae-Berothidae)] y retoman el criterio de que la familia representa un grupo hermano del resto de los neurópteros
del que divergió hace c. 294 ma durante finales del Carbonífero-principios del Pérmico, diversificándose las dos principales
subfamilias actuales (Coniopteryginae, Aleuropteryginae) durante el Triásico.
Con unas 450 especies descritas (Meinander, 1972a, 1990; Aspöck & Aspöck, 1999; H. Aspöck et al., 2001; New, 2001b) y numerosas sinonimias propuestas, unas 500 especies actuales son conocidas (Aspöck & Aspöck, 2007). Se desconoce el margen de variabilidad de muchas especies, y por el contrario, especies de muy amplia distribución presentan
una marcada variabilidad que hacen difícil definir inequívocamente diferentes taxa (p. ej.: Semidalis spp.), y autores como Tauber & Adams (1990) sugieren que el número de especies (sibling species) puede incrementarse hasta un 35 % cuando sean tenido en cuenta otros
elementos (microhábitats, morfología larvaria, etc.), y probablemente aún mayor será el número cuando sean estudiados a partir
de las nuevas herramientas moleculares.
En el Paleártico Occidental está representada por 108 especies pertenecientes a 11 géneros, y en Europa son conocidas 47 especies
de 9 géneros de esta familia (H. Aspöck et al., 1980, 2001; Aspöck et al., 2015), a las que recientemente se ha añadido una nueva especie (Helicoconis tatrica descrita de Eslovaquia como Helicoconis tatricus por Vidlička, 2014, el género Helicoconis es femenino, y si el autor quería referirse a las Montañas Tatra de donde fue descrita, como accidente geográfico, y no un
país, debería haber empleado la terminación-ensis) de Eslovakia, a las que ahora añadimos nuevos elementos.
La Fauna Ibérico-Balear española posee (salvo Vartiana, género monoespecífico con V. necopinata Aspöck & Aspöck, 1965 conocida de Grecia, Anatolia y Líbano) representantes de todos los géneros de esta familia presentes
en la Fauna neuropterológica Europea, y como ocurre en muchos otros órdenes de insectos, es particularmente rica en especies
de neurópteros, ya que nada menos que 197 especies están presentes en su fauna: Megaloptera (3 spp.), Raphidioptera (16 spp.)
y Planipennia o Neuroptera s. str. (178 spp.), representando en varias familias entre el 100 % a más del 75 % del total de especies europeas conocidas, y en
el caso de la familia que tratamos incluye más del 81 % de las especies europeas, con 39 especies en la fauna ibérica, de
las 48 especies válidas hasta hoy citadas en Europa (H. Aspöck et al., 1980, 2001; H. Aspöck, 1992; Aspöck & Hölzel, 1996; Aspöck & Aspöck, 1999; Monserrat, 2011; Monserrat & Triviño, 2013; U. Aspöck et al., 2015), algunas endémicas de nuestra fauna, y otras únicamente citadas de la Península Ibérica dentro del Continente Europeo. La
mayoría de la información corresponde a la zona peninsular española, pero también muchas de las especies ibéricas están citadas
de Portugal continental, de las que Letardi et al. (2013) listan 14 especies en esta parte ibérica de Portugal. Obviamente estos porcentajes son relativos, ya que son varias las especies
citadas en nuestra fauna que requieren confirmación, otras especies europeas han sido últimamente ratificadas como especies
válidas o han sido recientemente sinonimizadas o descritas, y otras están en fase de discusión y/o aceptación generalizada,
elementos que, en parte, pretendemos dilucidar mediante la presente contribución.
Debe indicarse que existen numerosas publicaciones relacionadas con la familia que tratamos, bien sobre fauna de otros continentes
o bien locales que escapan a la intención de esta contribución. Para los interesados en conocer información sobre sus subfamilias,
tribus, géneros, subgéneros y especies a nivel mundial se recomienda Oswald & Penny (1991) quienes listan los géneros conocidos y sus sinonimias, y Oswald (2013a) quien aporta toda la información sobre cada uno de sus géneros y especies descritas y Oswald (2013b) quien ofrece la información bibliográfica pertinente.
Sus especies alcanzan tamaños proporcionalmente pequeños, especialmente en relación a otras familias de neurópteros (la longitud
de alas anteriores rara vez supera los 5 mm, normalmente entre 1,8-5 mm), algo menores en los machos (Rousset, 1960a). Aunque su aspecto recuerda superficialmente a algunos homópteros sobre los que predan, especialmente a los aleyródidos,
por su tamaño, aspecto y su cuerpo blanquecino cubierto de ceras, son de morfología inconfundible (Figs. 1-9). Normalmente poseen el tegumento pardo muy oscuro y especialmente esclerificado (particularmente la cabeza, tórax y parte
de la genitalia), aunque poco después de nacer ya son de coloración blanca o blancuzca pulverulenta, debido a estas secreciones
céreas que producen unas glándulas tegumentarias que más tarde citaremos. Ocasionalmente poseen manchas oscuras en las membranas
de las alas, especialmente en las anteriores (Figs. 85, 88), y en muchas especies tropicales pueden tener el tegumento, y especialmente las antenas, con áreas o partes de tonos alternos
pardo claro/oscuro.
Dejando al margen las subfamilias y géneros no europeos, de los que sólo haremos alguna referencia cuando creamos conveniente,
y circunscribiéndonos a las especies de nuestras latitudes, pasamos a anotar los principales caracteres diagnósticos de esta
familia (Figs. 1-12).
|
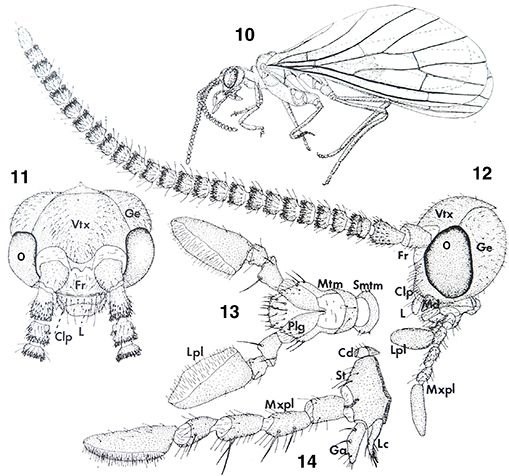 Figs. 10-14.— Morfología del imago, 10: aspecto general del imago, 11-14: Coniopteryx tineiformis (♂), 11: cabeza, vista frontal, 12: ídem, vista lateral, 13: labio, vista ventral, 14: maxila, vista ventral (Cd: cardo,
Clp: clípeo, Fr: frente, Ga: galea, Ge: gena, L: labio, Lc: lacinia, Lpl: palpo labial, Md: mandíbula, Mtm: mentum, Mxpl:
palpo maxilar, O: ojo, Plg: palpiger, Smtm: submentum, St: estipe, Vtx: vértex). A diferentes escalas. Adaptado de Meinander
(1972a). Figs. 10-14.— Morfología del imago, 10: aspecto general del imago, 11-14: Coniopteryx tineiformis (♂), 11: cabeza, vista frontal, 12: ídem, vista lateral, 13: labio, vista ventral, 14: maxila, vista ventral (Cd: cardo,
Clp: clípeo, Fr: frente, Ga: galea, Ge: gena, L: labio, Lc: lacinia, Lpl: palpo labial, Md: mandíbula, Mtm: mentum, Mxpl:
palpo maxilar, O: ojo, Plg: palpiger, Smtm: submentum, St: estipe, Vtx: vértex). A diferentes escalas. Adaptado de Meinander
(1972a).
Figs. 10-14.— Imago Morphology, 10: general aspect of imago, 11-14: Coniopteryx tineiformis (♂), 11: head, frontal view, 12: ditto, lateral view, 13: labium, ventral view, 14: maxilla, ventral view (Cd: cardo, Clp:
clypeus, Fr: frons, Ga: galea, Ge: gena, L: labium, Lc: lacinia, Lpl: labial palpus, Md: mandible, Mtm: mentum, Mxpl: maxillary
palpus, O: eye, Plg: palpiger, Smtm: submentum, St: stipes, Vtx: vertex). At different scales. Adapted from Meinander (1972a).
|
|
La cabeza es pequeña, especialmente en Brucheiserinae, hipognata, con dos grandes ojos compuestos bien desarrollados, traslúcidos,
con un diafragma interno negro, y ampliamente lateralizados, ocasionalmente con dimorfismo sexual (Adams, 1973) y sin ocelos presentes (Figs. 10-12). Está fuertemente esclerificada y posee sutura temporal bien marcada. Son varios los géneros en los que machos, hembras
o ambos poseen áreas membranosas en el vértex (Figs. 16, 20) o estructuras en el vértex y en la frente (Figs. 15, 17, 229, 240) (Ohm, 1965; Meinander, 1969a, 1972a, 1981, 1983b, 1998b; Johnson, 1976; Monserrat, 1989a, 2002, 2005a, 2005b; Sziráki, 1997, 2004, 2009; Zimmermann, 2005a, 2005b, etc.), acompañando a otros elementos cefálicos con dimorfismo sexual (Coniopteryx, Semidalis, Neosemidalis, Heteroconis; Pampoconis, Aleuropteryx, Helicoconis, Bidesmida) que complementan la morfología cefálica habitual existente y que probablemente intervengan en la captación de feromonas
en el reconocimiento y la atracción sexual, como veremos al hablar de su biología, así como ayuda en el apareamiento y cópula
(Tjeder, 1957; Meinander, 1969a, 1972a, 1981; Murphy & Lee, 1971; Monserrat, 1982c, 1989a, 1995, 2002; Zimmermann, 2005a, 2005b).
Las antenas son moniliformes, generalmente bastante más cortas que las alas anteriores, algo más largas en Brucheiserinae.
El número de flagelómeros es generalmente variable (16-60) (hasta más de 70 flagelómeros en Flintoconis), en ocasiones es más fijo (Heteroconis), y aunque se ha citado cierta variabilidad en su número (Killington, 1936), con frecuencia ha sido utilizado en la diferenciación de algunas especies (Conwentzia). Son generalmente pardos, aunque en especies tropicales muy frecuentemente con alternancia de colores formando bandas más
claras u oscuras (Meinander, 1972a; Monserrat & Díaz-Aranda, 1988b). El escapo es mayor que el pedicelo (Figs. 11, 12, 21, 94), a veces es algo globoso (Coniopteryx) o es más alargado (Helicoconis, Heteroconis, Semidalis). El pedicelo generalmente es alargado, a veces globoso y/o con setas escuamiformes en los machos (Coniopteryx) (Figs. 12, 19, 21, 22, 238, 239), o con estructuras que favorecen la cópula (Aleuropteryx) (Fig. 94). Flagelómeros moniliformes, con frecuente dimorfismo sexual en el número, la longitud y en la forma de algunos segmentos,
a veces con flagelómeros semifusionados (Tjeder, 1931), normalmente son más largos y estrechos en las hembras que en los machos, en los que es frecuente cierta setación especializada,
bien con anillos de setas escuamiformes (Coniopteryx, Neosemidalis) (Figs. 12, 180, 194, 211), a veces con setas muy desarrolladas y/o ganchudas o diversas estructuras sobre algunos flagelómeros (Heteroconis, Coniopteryx, Neosemidalis) (Karny, 1923; Meinander, 1969a, 1972a, 1981, 1990; Murphy & Lee, 1971; Sziráki, 2004, 2015) que sin duda favorecen el apareamiento y la cópula.
Las piezas bucales son de tipo masticador (Figs. 11-14, 20). El clípeo es pequeño pero con frecuentes setas. Las mandíbulas son pequeñas, simétricas y portadoras de un dentículo interno, maxilas bien desarrolladas, con galea unisegmentada (Coniopteryginae) o trisegmentada (Aleuropteryginae), lacinia muy pilosa,
palpo maxilar formado por 5 artejos, en ocasiones muy modificados en los machos (Coniopteryx neárticos) y palpo labial de 3 artejos (Figs. 12-14) (Enderlein, 1906; Killington, 1936; Tjeder, 1957; Meinander, 1972a; New, 1989; Stelzl, 1992). En algunos géneros (Semidalis) se ha citado dimorfismo sexual en las piezas bucales (Zimmermann 2005b).
El tórax compacto, poco piloso, de similar color que el resto del tegumento, aunque frecuentemente porta manchas circulares
más oscuras en la región tergal del pterotótax. Protórax corto, con pronoto transverso, generalmente menos esclerificado que
el pterotórax (excepto en especies braquípteras o ápteras), el cual suele estar más desarrollado (Figs. 1, 2, 10). Tanto el tórax como las alas están especialmente cubiertos por la citada secreción cérea de las glándulas hipodérmicas
tegumentarias que le dan el color blanco pulverulento característico (Fig. 2).
|
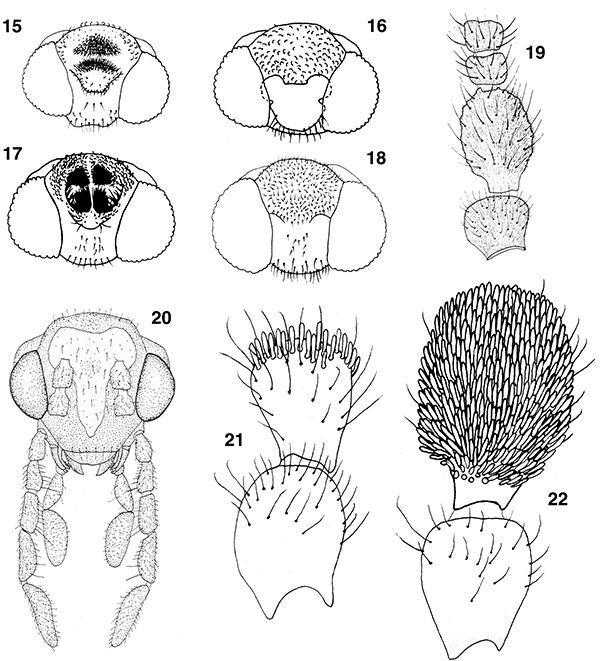 Figs. 15-22.— Morfología del imago, 15-18: esquema de la cabeza masculina de Coniopteryx spp., mostrando sus elementos sobre el vértex, de: 15: C. perisi, 16: C. kerzhneri, 17: C. sp., 18: C. atlasensis, de Monserrat (2002), 19: primeros segmentos antenales del macho de C. perisi, de Monserrat (1976a), 20: Helicoconis lutea, cabeza, vista frontal, de Meinander (1972a), 21: escapo y pedicelo en el macho de C. atlasensis, 22: ídem de C. perisi, de Monserrat (2002). A diferentes escalas. Figs. 15-22.— Morfología del imago, 15-18: esquema de la cabeza masculina de Coniopteryx spp., mostrando sus elementos sobre el vértex, de: 15: C. perisi, 16: C. kerzhneri, 17: C. sp., 18: C. atlasensis, de Monserrat (2002), 19: primeros segmentos antenales del macho de C. perisi, de Monserrat (1976a), 20: Helicoconis lutea, cabeza, vista frontal, de Meinander (1972a), 21: escapo y pedicelo en el macho de C. atlasensis, 22: ídem de C. perisi, de Monserrat (2002). A diferentes escalas.
Figs. 15-22.— Imago Morphology, 15-18: scheme of male head in Coniopteryx spp., showing its elements on vertex, of: 15: C. perisi, 16: C. kerzhneri, 17: C. sp., 18: C. atlasensis, from Monserrat (2002), 19: first antennal segments in male of C. perisi, from Monserrat (1976a), 20: Helicoconis lutea, head, frontal view, from Meinander (1972a), 21: scape and pedicel in male of C. atlasensis, 22: ditto of C. perisi, from Monserrat (2002). At different scales.
|
|
Las alas están bien desarrolladas, son subiguales, amplias y generalmente de ápices redondeados, algo más largas las anteriores
(Figs. 1, 2, 10, 23, 24, 160, 243), aunque son de pequeño tamaño, de mucho menor tamaño que en otras familias de neurópteros (longitud de las alas anteriores
en especies de nuestras latitudes: 1,8-5 mm). y raramente superan los 5 mm (longitud de las anteriores en Conwentzia hasta 8,3 mm, en Helicoconis o Flintoconis hasta 9,2 mm), son ovoides, a veces algo más alargadas (Flintoconis, Brucheiser, Cryptoscenea), y ocasionalmente hay géneros con alas algo curvadas (Coniocompsa, Brucheiser, Cryptoscenea), a veces falcadas (Heteroconis) o con reducción de las alas posteriores (Conwentzia, Coniopteryx) (Fig. 90), y en elevadas latitudes, poblaciones isleñas o de alta montaña también hay reducción en las alas posteriores con ejemplares
macrópteros, braquípteros, micrópteros o dimórficos, incluso ápteros (Helicoconis, Coniopteryx) y/o algo más esclerificadas y coriáceas, especialmente la venación y el margen (Brucheiser) (Perkins, 1899; Tullgren, 1906; Withycombe, 1922a, 1922b, 1924a, 1924b, 1925a, 1925b; Navás, 1927a; Killington, 1936; Kimmins, 1950; Tjeder, 1960, 1972; Messner, 1965; Meinander, 1971, 1972a, 1974d, 1990; Riek, 1975; New, 1986, 1989, 2003; Monserrat, 2002; Tauber et al., 2009). Habitualmente, en estado de reposo las mantienen plegadas “en tejadillo” (tectiformes) sobre el abdomen (Figs. 1, 2, 10), hecho habitual en casi todas las familias de Neuropteroidea (excepción en Psychopsidae y en algunos géneros y/o especies
de mirmeleónidos, crisópidos o hemeróbidos, ejs.: Tricholeon, Solter, Ankylopteryx, Chrysoperla, Psectra, Notiobiella, etc.). Mayoritariamente la membrana alar es hialina, levemente coloreadas de gris, a veces de tonos más pardo-grisáceo (Conwentzia), a veces marcadamente intenso (Parasemidalis fuscipennis). Muchas especies, en particular de géneros tropicales (Heteroconis, Coniocompsa, Cryptoscenea, Helicoconis, Spiloconis, Neoconis, Pampoconis, Nimboa, Incasemidalis, Hemisemidalis,
Semidalis), inclusive ya en especies fósiles (Meinander, 1972a; Monserrat & Díaz-Aranda, 1988b; Nel et al., 2005), y alguna de las nuestras poseen manchas oscuras sobre la membrana, bien aisladas de forma llamativa (Aleuropteryx maculata, A. minuta, Nimboa adelae) (Figs. 85, 88, 104, 112, 243) o como sombras más oscuras sobre el margen anterior o en las celdillas marginales (Aleuropteryx juniperi) (Fig. 1) (probablemente las manchas negras sobre la membrana de las alas en estas especies pueda actuar como elemento mimético con
ciertos homópteros para confundirse, aún más, con ellos), pero las alas siempre están cubiertas de una secreción cérea segregada
por glándulas hipodérmicas que les da una coloración externa blanca intensa y probablemente disminuya su peso específico relativo
favoreciendo el vuelo. El perostigma es imperceptible en ambas alas (Figs. 23, 24). La venación está marcadamente reducida, muy simplificada y similar en ambas alas y con venillas transversales escasas (Figs. 10, 23, 24, 160, 250). En las alas anteriores el campo costal no porta más de dos venillas que carecen de bifurcación o de vena humeral recurrente,
la vena Sc bifurcada en el extremo (echo inusual en el orden y que comparten con Megaloptera), el único SR surge del R cerca
de la región media de las alas, R y M fusionadas en su base, y las venas longitudinales nunca se bifurcan varias veces, ni
cerca del margen alar, y dos venas anales son frecuentes (Figs. 10, 23, 24, 160, 250). La venación está aún más reducida en las porteriores, el único SR surge del R en la región basal (Aleuropteryginae) o cerca
de la región media de las alas (Coniopteryginae) y dos venas anales son frecuentes (Figs. 23, 24). En Brucheiserinae la venación es de tipo reticulado y muy anómala (Brucheiser) o alargadas y venación subparalela con abundantes venillas trasversales (Flintoconis) (Navás, 1927a; Riek, 1975; New, 2001b; Sziráki, 2007a), que desde nuestro punto de vista poco tienen que ver entre sí para definir la subfamilia ni la pertenencia de Flintoconis a esta misma subfamilia. La venación puede estar sometida a un cierto grado de variabilidad, incluso dentro de la misma especie
(Houser, 1914; Withycombe, 1922a; Tjeder, 1931; Rousset, 1960a; Zelený, 1961a). Venación en ambas subfamilias de nuestra fauna según las figuras 23 y 24. Tricosoros (pequeños espesamientos sobre la vena
costal en el extremo de las bifurcaciones de las venas longitudinales) ausentes (Figs. 10, 23, 24, 160, 217), pero las setas marginales están generalmente bien desarrolladas y son abundantes, especialmente en el margen posterior
de las alas anteriores, y abundan las macrotrichias sobre las venas longitudinales (destacan dos grandes setas sobre M en
Aleuropteryginae, Figs. 23, 93, 104, 112), también microtrichias abundan sobre toda la membrana, y carecen de nygmata (pequeños puntos cuticulares oscuros que pueden
aparecer en las alas anteriores, posteriores, o en ambas en algunas familias de neurópteros). Aparato de acoplamiento entre
ambas alas poco desarrollado (Figs. 23, 24), al máximo algunas setas a modo de corto hámuli en la vena costal de las alas posteriores y ciertas setas unciformes en
el lóbulo yugal de las anteriores (New, 1989).
|
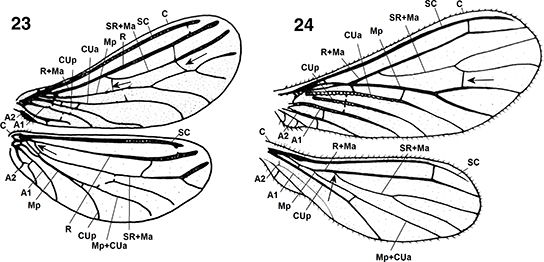 Figs. 23-24.— Terminología alar en: 23: Aleuropteryginae: Aleuropteryx iberica, 24: Coniopteryginae: Coniopteryx ezequi (A: anal, C: costal, CUa: cubital anterior, CUp: cubital posterior, Ma: mediana anterior, Mp: mediana posterior, R: radio,
SC: subcostal, SR: sector del radio). Las flechas señalan algunos caracteres usados en las claves. A diferentes escalas. 23
de Monserrat (1977b), 24 de Monserrat (1984g). Figs. 23-24.— Terminología alar en: 23: Aleuropteryginae: Aleuropteryx iberica, 24: Coniopteryginae: Coniopteryx ezequi (A: anal, C: costal, CUa: cubital anterior, CUp: cubital posterior, Ma: mediana anterior, Mp: mediana posterior, R: radio,
SC: subcostal, SR: sector del radio). Las flechas señalan algunos caracteres usados en las claves. A diferentes escalas. 23
de Monserrat (1977b), 24 de Monserrat (1984g).
Figs. 23-24.— Wings Terminology in: 23: Aleuropteryginae: Aleuropteryx iberica, 24: Coniopteryginae: Coniopteryx ezequi (A: anal, C: costal, CUa: anterior cubital, CUp: posterior cubital, Ma: anterior median, Mp: posterior median, R: radius,
SC: subcostal, SR: radial sector). At different scales. 23 from Monserrat (1977b), 24 from Monserrat (1984g).
|
|
Las patas son marchadoras, algo más desarrolladas las anteriores (Fig. 10), a veces las coxas son libres y a veces muy pilosas, los fémures son cilíndricos, en ocasiones los machos con espinas en
la cara interna (Fig. 179) (Coniopteryx, Semidalis) (Tullgren, 1906; Enderlein, 1906; Mjöberg, 1909; Tjeder, 1931; Murphy & Lee, 1971; Sziráki, 1997, etc.) y las tibias están comprimidas lateralmente y ensanchadas en su zona media, las posteriores son algo más largas (Fig. 10). Tarsos de 5 tarsómeros, más largo el basal, en Brucheiserinae el cuarto tarsómero está especialmente bilobulado, semejante
a lo que hallamos en Megaloptera: Sialidae (también evidente en otros géneros como Helicoconis o Conwentzia), uñas tarsales curvas y simples, empodio pequeño y apuntado, y arolio ausente. Estos caracteres ya están definidos en ejemplares
fósiles desde el Cretácico Superior (Perrichot et al., 2014).
El abdomen es algo más corto que sus alas (Figs. 1, 10) y está más escasamente esclerificado que los anteriores tagmas, pues posee amplias zonas tergales membranosas, y también
está externamente cubierto por las secreciones céreas citadas. Formado por diez segmentos de los que estructuras del 8° y/o
9° se incorporan a la terminalia y genitalia, por lo que solo los más anteriores se aprecian claramente separados, con tergos
y esternos bien definidos y amplias y extensibles áreas pleurales (Figs. 95, 96, 114, 213), con estigmas en los segmentos abdominales 1-8 en Brucheiserinae y Coniopteryginae, 1-7 y Aleuropteryginae (la pérdida de
estigma en el último segmento en Coniopteryginae ha sido refutada por Zimmermann, 2005b y Zimmermann et al., 2009, donde un 8° estigma ha sido también descrito).
Son características las glándulas de la cera, que se abren formando hileras de pequeños puntitos por donde se segregan las
sustancias céreas en diferente posición y presentes en ambos sexos (Figs. 95, 96, 108, 289, 291), en Coniopteryginae más frecuentes en las zonas tergales y laterales de los segmentos torácicos y abdominales, en la región
basal de las alas y también están presentes en el vértex y el escapo, y están especialmente desarrolladas en Aleuropteryginae,
formando hileras sobre algunos esternitos y terguitos abdominales, y en Brucheiserinae están dispuestas en forma de arco sobre
los esternitos de los segmentos 2°-6° en las hembras y 2°-7° en los machos. Estos pequeños orificios tegumentarios se abren
sobre el tegumento en forma de roseta, y es donde desembocan las glándulas hipodérmicas que segregan la cera, mayoritariamente
formada por ácidos grasos, hidrocarburos y alcoholes (para su composición se recomienda Nelson et al., 2003), cera que los cubre completamente (excepto ojos y el flagelo de antenas), dándoles un aspecto similar a lo que hallamos
en aleyródidos y cóccidos (Fig. 2), hecho que no nos parece casual, sino de forma convergente, ya que habitualmente sobre ellos predan (esta posibilidad ya
había sido sugerida por Penny et al., 2007), y les da su color y aspecto característico. Estas ceras están segregadas en forma de microscópicos recurvados filamentos,
y acaban por desprenderse y los ejemplares las extienden activamente sobre su cuerpo con las tibias y fémures de sus patas
anteriores y posteriores poco después de emerger el imago (Withycombe, 1922a, 1923, 1925a; Killington, 1929, 1936; Tjeder, 1957; Meinander, 1972a; Navone, 1987; Nelson et al., 2003).
También son curiosas y características de esta familia las llamadas “plicaturas”, ausentes en Coniopteryginae y especialmente
desarrolladas en Aleuropteryginae (particularmente en los segmentos III-V, menos en VI y VII), y en Brucheiserinae, en posición
lateral sobre los esternitos distales del abdomen (Fig. 96), con orificios glandulares próximos o alrededor de ellas (Zimmermann, 2005b; Zimmermann et al., 2009) (Fig. 96). Estas estructuras circulares, lenticulares u ovoides y membranosas, se han citado en ocasiones (Enderlein, 1906; Lacroix, 1924; Withycombe, 1925a; Killington, 1936; Sziráki, 2007b; Zimmermann et al., 2009) como secretoras de sustancias repugnatorias o feromonas, como respiratorias, como captadoras de humedad ambiental, sensoriales,
o como “evaginables” por presión de la hemolinfa y con musculo retractor, como ocurre con el tubo ventral de los colémbolos,
aunque este último hecho (“evaginables/evaginadas”) nunca lo hemos observado en ejemplares vivos y, que sepamos, solo se conoce
de forma aparente en formas fósiles (†Archiconis electrica) y pocas veces en las especies actuales, donde estas formaciones membranosas evaginables, de aspecto triangular/cuadrangular,
parecen permanecer en ciertos géneros (Coniocompsa, Helicoconis, Spiloconis), y son de función desconocida, aunque han sido asociadas a las vesiculas o tubos ventrales evaginables de algunos apterigotas
(Ejs.: Collembola, Microcoryphia, Zygentoma, Protura) (Enderlein, 1906; Carpenter, 1955; Tjeder, 1957, 1960; Meinander, 1972a; New, 1989; Zimmermann et al., 2009). Otras estructuras membranosas evaginables en el abdomen (pleuritocavae, glándulas evaginables, gonocoxitos), de difíciles
holologías con ellas, están presentes en otras familias de neurópteros (Chrysopidae, Mantispidae, Nemopteridae, Crocinae,
Osmylidae, Nevrorthidae) o megalópteros (Sialidae, Corydalidae).
Los segmentos distales del abdomen están sexualmente muy especializados en los ♂♂, sin callo cercal ni tricobotrias presentes
y con estructuras genitales externas e internas bien desarrolladas en ellos, y con elementos característicos que se exponen
en cada género. Menos aparentes, escasamente esclerificados externamente y vias genitales internas muy específicas en las
♀♀, a veces con placas subgenitales y espermatecas esclerificadas, y siempre ausencia de ovopositor (Figs. 96, 102, 103, 105-108, 113-117, 133-138, 254, 255, 258, 259, 264-266, 289-315).
La genitalia masculina y femenina es muy variable según las subfamilias, géneros, subgéneros y especies, siendo de homologías
difíciles en relación entre los diversos géneros y entre esta familia y otras familias de neurópteros (Tjeder, 1954, 1956, 1957, 1970; Acker, 1960; Tuxen, 1970; Meinander, 1972a). El segmento abdominal 11° está obliterado, por lo que carecen de cercos y/o de ovopositores, y tampoco portan callo cercal
sensorial, característico de muchas otras familias de neurópteros. Como hemos indicado, los machos presentan pene y, curiosamente
y diferente a los restantes neurópteros, poseen los testículos solo funcionales en las fases larvarias, mientras que en los
imagos el semen se acumula en un reservorio ventral (vesícula seminal llena de gametos) que finaliza a través de un ducto
eyaculador (Figs. 100, 101, 166, 184, 187, 216, 223, 235, 249, 260), con su propia musculatura, en el pene (Withycombe, 1925a; Stitz, 1931; Killington, 1936; Meinander, 1972a; New, 1989), por lo que la inseminación es interna y sin concurso de espermatóforo, estructura presente en muchas otras familias de
neurópteros (Megaloptera, Raphidioptera, Neuroptera: Osmylidae, Sisyridae, Berothidae, Mantispidae, Hemerobiidae, Chrysopidae,
etc.), y que representa uno de los elementos más evolucionados dentro de los Neuropteroidea. En las hembras la espermateca
o bolsa copuladora, frecuentemente esclerificada, recoge el esperma hasta el momento de la fecundación de los óvulos (Figs.
96, 102, 103, 105-108, 113-117, 133-138, 254, 255, 258, 259, 264-266, 289-315).
Aunque otros elementos de biología, distribución, morfología externa y pigmentación tegumentaria y alar pueden ayudar, el
estudio de la genitalia (interna y externa) es imprescindible para la correcta identificación de las especies de esta familia,
y en las que nos competen, tanto la genitalia masculina como femenina de las especies ibéricas se anotan a lo largo del texto
en cada caso. Se sigue la terminología general comúnmente aceptada de Meinander (1972a) y Aspöck et al. (1980), aunque es relativamente frecuente que, también en esta familia, diversos autores hayan introducido cambios en la
terminología clásica, en función de sus interpretaciones.
A pesar de su interés en el terreno de la Entomología Aplicada, sus fases larvarias han despertado menor atención que en otras
familias, especialmente Chrysopidae (Withycombe, 1924b; Silvestri, 1942; New, 2001a), y solo existen datos para menos de la tercera parte de los géneros conocidos. Aunque nos detendremos en sus fases juveniles
al hablar de su biología, adelantemos que su larva es depredadora de huevos y larvas de insectos y de pequeños artrópodos
como ácaros, homópteros y otros insectos, etc., y es de tipo campodeiforme y de vida libre (Figs. 5-7, 44, 53, 65, 71). La fase larvaria madura que se dispone a pupar, tras tejer el capullo (Figs. 8, 76), adopta una posición quiescente y curvada antes de transformarse en pupa (Figs. 9, 77). La pupa es activa, dectica y exarada (Figs. 78, 79).
Más información general o relativa a nuestra fauna en: Hagen, 1856; Dziędzielewicz, 1905; Enderlein, 1906; Roepke, 1917; Arrow, 1917; Withycombe, 1923; Meinander, 1972a, 1974a; Ripolles & Melia, 1980; Castellari, 1980; Aspöck & Aspöck, 1980, 1999, 2001; Mansell, 1986; Tauber & Adams, 1990; Tauber, 1991; Gepp, 1999; Sziráky & Flint, 2007.
DATOS GENERALES SOBRE SU DISTRIBUCIÓN, BIOLOGÍA, COMPORTAMIENTO Y CICLOS BIOLÓGICOS
Los coniopterígidos no son precisamente insectos “familiares”, como son las mariposas, las libélulas, las abejas, los escarabajos,
etc., incluso otros grupos de neurópteros más conocidos (las crisopas, las nemópteras, las hormigas-león, etc.), sino insectos
que suelen pasar desapercibidos en la Naturaleza, al tratarse de un grupo de pequeño tamaño, poco llamativos, de actividad
crepuscular-nocturna, y con escasa capacidad de vuelo.
Esta curiosa familia tiene una distribución cosmopolita, salvo la Antártida y zonas particularmente elevadas y frías, e incluye
especies mayoritariamente habitantes de zonas tropicales, subtropicales, y en menor medida en zonas templadas, generalmente
arboladas o boscosas, evitando en general medios más adversos, aunque en ocasiones hay especies adaptadas a medios con muy
escasa y aislada vegetación, a veces están presentes en medios térmicos, secos y espacios abiertos y soleados, y muchas especies
están adaptadas a medios marcadamente eremiales, xéricos y subdesérticos (Tjeder, 1957, 1987; Meinander, 1972a, 1977, 2002; New, 1986; Sziráki, 1992e, 1997; Monserrat, 1996b; Sziráki & Harten, 2006, etc.), y están ausentes en climas fríos, aunque se han citado en muy elevadas latitudes y altitudes, de hasta 3.300-3.886
m en zonas de alta montaña (Carpenter & Lestage, 1928; Stitz, 1928; Tjeder, 1940, 1943, 1972; Kimmins, 1950; Meinander, 1962a; Schedl, 1970; Meinander, 1972a, 1972c; Rausch et al., 1981; Greve & Kobro, 1998; Sziráki, 1999; Popov, 2002a, 2002b, etc.), y ahora citamos material recolectado a 2.070 m, 2.200 m, 2.500 m, 2.550 m, 2.580 m y 2.800 m entre nuestras especies.
Aunque pueden recolectarse mediante otros métodos (ver material citado y ejemplos tenemos en Vas et al., 2001 o Vidlička, 2015), con mucha frecuencia acuden a la luz (Killington, 1936; Lewis & Taylor, 1964; Ward, 1965; Andersen & Greve, 1975; New, 1986, 1989; Günther, 1991; Greve & Kobro, 1998; Ábrahám et al., 2003; Penny et al., 2007, etc.), tanto machos como hembras (aunque erróneamente Withycombe, 1924a, 1924b mencionaba que sólo lo hacían los machos), y algunas especies de biología poco conocida sólo se han recolectado por este
medio, y por esta fotofilia acuden accidental-ocasionalmente al interior de nuestras viviendas, y muchas especies son habituales
en nuestros parques y en espacios ajardinados (Withycombe, 1924b; Ward, 1965; Czechowska, 1982, 1986; Monserrat, 1985e, 1995; Barnard et al., 1986; Günther, 1991; Plant, 1992a, 1992b, 2001; Pantaleoni, 1995; Monserrat et al., 1999; Gruppe, 2007a; Valencia, 2009, etc.).
Imagos:
Como hemos indicado, los coniopterígidos son poco familiares debido a su pequeño tamaño y a que sus imagos son insectos mayoritariamente
de actividad crepuscular/nocturna (Castellari, 1980; New, 1986, 1989; Vas et al., 1999; Ábrahám & Mészáros, 2006), aunque se han citado en ocasiones especies con dos periodos de vuelo nocturno en los que son muy activas (Conwentzia psociformis), tanto tras el atardecer, como antes del amanecer (Lewis & Taylor, 1964), o solo desde un poco antes del atardecer, e incluso activas durante el día (Ábrahám & Vas, 1999; Monserrat, 2005b, y observaciones ahora citadas por el autor). Con respecto a estos elementos de su biología en el vuelo, comentemos que,
en general, sus imagos son muy activos, ágiles, y aunque son buenos voladores, sus vuelos en general son cortos y breves,
y por ello con escasa capacidad de dispersión, aunque han sido frecuentemente observados en vuelos sostenidos y pueden formar
enjambres al atardecer y/o ser arrastrados por el viento, en ocasiones a cierta altura, incluso mar adentro, y/o poseer, en
realidad, mayor capacidad de vuelo y de dispersión (Britten, 1916; Southwood, 1957; Murphy & Lee, 1971; Cheng & Birch, 1978; Sugg et al., 1994; Monserrat et al., 1999; Monserrat, 2005b, etc.), y de hecho las trampas de succión son un buen método para recolectarlos y Vas et al. (2001) lo demuestra. Curiosamente, algunas especies aumentan su actividad antes de una tormenta (Southwood, 1957), y parece comprobado que varias especies resisten bien los efectos de los fuertes vientos (Vidlička, 2015). Al margen del vuelo, sus alas son utilizadas como elemento de reconocimiento, apaciguamiento y atracción sexual durante
el cortejo, de cripsis para acercarse a ciertas presas de similar aspecto (especialmente aleyródidos) y en situaciones de
peligro, a veces saltan y se dejan caer al suelo y, como ha sido observado en otras familias de neurópteros (Hemerobiidae,
Sisyridae, Dilaridae, etc.), han sido observados adoptando por un breve tiempo un comportamiento de tanatosis con las alas
plegadas y así disuadir o mitigar el interés del potencial depredador (Withycombe, 1924b; Killington, 1936, 1937a), y este comportamiento lo hemos observado en multitud de ocasiones.
Los imagos de Coniopterygidae son depredadores generalistas de pequeños y/o muy pequeños artrópodos fitófagos, bien sésiles
o de movimientos lentos y tegumento blando, y también de sus puestas, especialmente homópteros: áfidos, aleyródidos, diaspídidos,
filoxéridos, pseudocócidos y cóccidos, también psíllidos, tisanópteros, huevos y larvas de otros insectos, etc., y en particular
de ácaros: Eriophyidae, Tetranychidae, en proporción correspondiente a su biología, morfología y tamaño. Por ello son excelentes
aliados de nuestros intereses contra estos insectos, Withycombe, 1923, 1924b; Balduf, 1939; Silvestri, 1942; Yang, 1951; Fleschner & Ricker, 1953; Collyer, 1964; Putman & Herne, 1966; Riek, 1975; New, 1975, 1986, 1989, 1991a; Sziráki, 1979; Stimmel, 1979; Castellari, 1980; Ripolles & Melia, 1980; García Marí et al., 1983; Stelzl, 1989, 1992; Dinkins et al., 1994; García Marí & González Zamora, 1999; Stelzl & Devetak, 1999; Canard, 2001; Gerling et al., 2001; McEwen et al., 2001; Nicoli Aldini, 2002; Soler et al., 2002; Miller et al., 2004; Pantaleoni, 2007; Monserrat & El-Serwy, 2010, entre otros muchos, comentan su interés aplicado en cultivos e intereses humanos, y de hecho se han introducido en numerosos
países algunas especies exóticas para su utilización como agentes de control de determinadas plagas [Ej: la oriental Spiloconis picticornis en cultivos de cítricos y aguacates en California (Badgley et al., 1955)], o han sido introducidas accidentalmente por acción humana en países o archipiélagos donde no son nativas [p. ej. Hawaii:
Zimmerman (1940, 1957), Nueva Zelanda: Kimmins & Wise (1962), Wise (1991, 1992) o Norteamérica: Meinander (1986)], para otros ejemplos ver, entre nuestras especies, la distribución de Aleuropteryx juniperi, Conwentzia psociformis o C. pineticola.
Sin duda la mayor o menor longitud de sus piezas bucales está en función de la capacidad de moción y protección de sus presas
(New, 1986). Killington (1936) anota las presas citadas entre las especies británicas y Canard (2001) anota una relación de presas observadas en diversas especies por diversos autores. Pueden llegar a ser extremadamente voraces
(Withycombe, 1923, 1924b, 1925a; Killington, 1936; Collyer, 1951; Castellari, 1980; Stelzl, 1991, etc.), habiéndose registrado, por citar algunos ejemplos, en Conwentzia pineticola, consumos de 30/40 ácaros en una hora (imagos) y de 15-20, 30-35 ácaros diarios (larvas) (Collyer, 1951), o Muma (1967) quien anota hasta 40 ácaros a la hora devorados por imagos de Semidalis vicina. Muchas especies complementan su dieta con elementos de alimentaciones más omnívoras, pues ingieren polen y néctar (Withycombe, 1923, 1924b; Lacroix, 1924; Narayanan, 1942; Fleschner & Ricker, 1953; Badgley et al., 1955; New, 1986, 1989; Stelzl, 1989, 1992; Monserrat, 2005b, etc.), pero al margen de lo anteriormente anotado, su régimen alimenticio mayoritario es depredador de pequeños artrópodos
de tegumento no demasiado esclerificado y de sus puestas (Figs. 5-7). No existe canibalismo, pero en ocasiones las ♀♀ en cautividad, particularmente si son vírgenes, devoran sus propios huevos
recién puestos. El aseo de sus piezas bucales tras la ingesta es obligatorio y frecuente (Collyer, 1951).
Sobre la elección de su hábitat, comentemos que, en general son bastante específicos en el sustrato vegetal sobre el que habitan
y se desarrollan, y por ello a veces son muy locales y normalmente poco frecuentes, aunque a veces pueden llegar a ser extremadamente
abundantes. En ocasiones algunas especies parecen ser verdaderamente estenotópicas (Aleuropteryx iberica, A. maculata, A. minuta, Coniopteryx parthenia, C. ezequi, C. kerzhneri, Parasemidalis triton, etc.), hecho que limita su distribución (amplitud de hábitat HA según Simpson, 1949 menor de 2: 1,1-1,9), y muestran una marcada especificidad en sus presas (estenofagia) que, a su vez, como ocurre normalmente
entre los fitófagos, se hallan sobre un tipo específico de vegetación, hecho que, entre otros elementos ambientales y siendo
insectos depredadores, les genera indirectamente una marcada especificidad en el tipo de plantas substrato sobre el que los
hallamos, sean coníferas, planifolios, vegetación herbácea, etc. Otras especies, por el contrario, son más euritópicas (HA
entre menor de 6: 2,0-5,2), dentro de una cierta amplia gama de plantas, si bien particulares de determinados ambientes, sean
de naturaleza mediterránea o eurosiberiana (Aleuropteryx loewii, Helicoconis interna, Conwentzia psociformis, C. pineticola, Coniopteryx esbenpeterseni, C. arcuata, C.
haematica, Semidalis pseudouncinata, S. vicina, Hemisemidalis pallida, etc.). Otras especies son declaradamente generalistas (Semidalis aleyrodiformis, Coniopteryx borealis) con amplitud de hábitat HA superior a 9: 9,3-12,8, y en ciertos casos desconocemos la vegetación específica sobre la que
se desarrollan y/o han sido mayoritariamente recolectadas a la luz (Semidalis pluriramosa, Coniopteryx ketiae, C. atlantica, Aleuropteryx remane, Helicoconis spp., Nimboa spp., etc.) y probablemente se desarrollen en vegetación herbácea o baja (Helicoconis, Xeroconiopteryx), mucho menos muestreada que los habituales fanerófitos o nanofanerófitos (Killington, 1936; Tjeder, 1957; Meinander, 1972a; New, 1986, 1989; Marín & Monserrat, 1987; Marín, 1994; Monserrat & Marín, 1992, 2001; Hölzel, 1998).
Se ha demostrado una estratificación de las diferentes especies en función de la altura del dosel arbóreo/vegetal, incluso
en plantas cultivadas (Murphy & Lee, 1971; Principi & Canard, 1974; Nielsen, 1977; Meinander & Penny, 1982; New, 1986, 1989; Hollier & Belshaw, 1993; Duelli et al., 2002; Gruppe, 2007a, etc.), y aunque mayoritariamente habitan en vegetación arbórea o arbustiva (Marín & Monserrat, 1987; Monserrat & Marín, 1992, 2001; Marín, 1994; Pantaleoni, 1996), también otras especies prefieren vegetación herbácea, o se han encontrado asociadas a medios marcadamente edáficos, musgos,
e incluso en el interior de galerías de embiópteros (Kimmins, 1950; Messner, 1965; Meinander, 1972a; Aspöck et al., 1980; New, 1986).
A veces los imagos se encuentran próximos a zonas infestadas por homópteros, donde pueden verse atraidos por sus potenciales
presas, y con frecuencia muestran señales de agregación (Southwood, 1957; Meinander, 2002; Aspöck & Aspöck, 2014), y varios ejemplares pueden ser recolectados en una misma rama o árbol (hasta cientos de ejemplares alrededor de un único
árbol cita Monserrat, 2005b en Semidalis kolbei, una especie neotropical), quizás también por atracción sexual o agregación en zonas específicamente elegidas para efectuar
bien el cortejo o las puestas (Withycombe, 1922a, 1924b; Killington, 1929; Johnson & Morrison, 1979), de hecho se han citado comportamientos de agregación sexual y se han observado hasta 15 machos revoloteando a menos de
medio metro alrededor de una hembra, de lo que se deduce la emisión de feromonas femeninas (Johnson & Morrison, 1979; Monserrat, 2005b), según ahora citaremos.
Su longevidad como imagos parece ser mayor en las hembras que en los machos, y quizás por eso parecen ser más frecuentes,
y según los datos oscila entre 3-122 días. Como ejemplos citemos a Collyer (1951) quien anota una vida media de 4 semanas en los machos y de 8 semanas en las hembras de Conwentzia en condiciones de laboratorio, Withycombe (1924b) quien anota longevidades de un mes en hembras de Conwentzia, Fleschner & Ricker (1953) anotan longevidades de 3-6, 19-76 y hasta 39-122 días en función de la variedad de dietas ofrecidas, Badgley et al. (1955) quienes anotan longevidades de 49-63 días en imagos de Spiloconis, y ciclos vitales completos desde la puesta de huevos a muerte de imagos de 92-131 días (110 días como media), Muma (1967) anota ciclos con una media de entre 35-43 días en función del tipo de alimento en Semidalis vicina y Castellari (1980) anota entre 14-32 días para Coniopteryx, con algunas diferencias en mayor longevidad en las hembras. Todos estos datos apuntan altas longevidades medias para ser
insectos tan pequeños.
Sobre su biología reproductiva Badgley et al. (1955) anotan, desde la emergencia del imago, un periodo mínimo de 3-5 días (hasta 10 días) para que maduren las gónadas femeninas
e iniciar el periodo reproductor, pero curiosamente, en la bibliografía europea no existen muchos datos sobre el comportamiento
de los individuos durante el cortejo y la cópula, y no parece fácil realizar observaciones en condiciones de laboratorio (Johnson & Morrison, 1979). Existen algunas observaciones muy interesantes e ilustrativas, así Withycombe (1922a, 1923) anota en Parasemidalis que el macho se aproxima a la hembra desde atrás e introduce su cabeza entre sus alas bajo su abdomen y se aferra a sus coxas
posteriores con sus mandíbulas y a sus patas posteriores y medias con sus patas anteriores, doblando su abdomen hacia arriba
para iniciar la cópula, conforme la hembra abre unos 45° sus alas y eleva el abdomen, mientras mantiene sus antenas proyectadas
horizontalmente, y también Collyer (1951) anota similar comportamiento en Conwentzia. Otras observaciones anotan acercamientos laterales o dorsales, y cópulas con cuerpos acoplados y en dirección cefálica opuesta
(Henry, 1976). Durante este proceso, las antenas del macho se abren casi 90°, algo menos las alas. A veces, si el macho no es aceptado,
la hembra vuela o camina rápidamente alejándose. Si el proceso de la cópula progresa, durante ella la hembra no suele permanecer
en reposo y el macho puede ser arrastrado por la hembra (Aspöck & Aspöck, 1999; Zimmermann, 2005b; Monserrat, 2005b), que puede desplazarse utilizando únicamente sus cuatro patas anteriores. La cópula dura entre 5-10, 10-15, 15-30 minutos,
según observaciones. Badgley et al. (1955) observan que, ocasionalmente, una misma hembra puede efectuar varias cópulas a lo largo de su vida, y puntualmente se ha
observado oofagia. A veces otros machos (hasta dos más) pueden acercarse a la pareja en cópula, incluso intentan “participar”
con el macho en cópula adoptando con él similar comportamiento copulador, aferrándose a sus patas, y hasta un tercer macho
puede intentar lo mismo con este último solícito visitante. En condiciones experimentales se han observado intentos de cópula
entre especies de géneros distintos, o entre especies diferentes del mismo género (Sziráki, 2002a), incluso con otros machos, y también se han observado hasta 15 machos revoloteando a menos de medio metro alrededor de una
hembra, de lo que se deduce la emisión de feromonas femeninas. No se han observado comportamientos postcopulatorios (Johnson
& Morrison, 1979). Más información en Withycombe, 1922a, 1923, 1925a; Lacroix, 1924; Stitz, 1931; Killington, 1936; Eglin, 1940; Narayanan, 1942; Collyer, 1951; Badgley et al., 1955; Muma, 1967, 1971; Yang, 1974; Henry, 1976, Johnson & Morrison, 1979; Castellari, 1980 y Monserrat et al., 2001.
En relación a otros comportamientos reproductores, se han observado vibraciones de las alas en ambos sexos como llamadas de
cortejo a intervalos de 1-2 segundos, durante cerca de una hora, mientras la hembra permanece en el sustrato o camina sobre
él, elevando y rotando su abdomen, similar comportamiento se ha observado en los machos (Johnson & Morrison, 1979), consecuentemente se han detectado tremulación/vibraciones en las alas como comportamiento inmediatamente previo a la cópula
y órganos sensoriales (escolopales, cordotonales y subgenuales) de captación de vibraciones (rango de frecuencia de 300-2.000
Hz) en la región media de las tibias de todas las patas (Johnson & Morrison, 1979; Devetak, 1998). Las vibraciones se trasmiten bien vía aire o a través del sustrato. Citemos que la emisión-recepción de sonidos es extremadamente
importante en la localización y selección sexual entre individuos de muchas familias de neurópteros, y ha sido especialmente
estudiado en Chrysopidae, con estructuras alares con órganos timpánicos sobre espesamientos en la base de algunas venas, y/o
con formaciones alares, torácicas (metanoto) y abdominales (esternitos) con funciones estridulatorias, y sin embargo estas
estructuras parecen poco desarrolladas en esta familia (Riek, 1967), pero aún así la comunicación acústica, también en esta familia, debe realizar una importante misión en el cortejo. Al margen
de las particularidades anatómicas citadas anteriormente en esta familia, recordemos que la trasferencia de esperma es directa,
merced al concurso de un pene en los machos, y no a través de ningún tipo de espermatóforo (Withycombe, 1925a).
Tras su emergencia como imago y tras las fases de cortejo y cópula, las hembras inician la fase de la puesta. La puesta de
huevos comienza, en condiciones experimentales, desde pocas horas a varios días, pudiéndose prolongar durante unos pocos días
a semanas, según las observaciones, incluso hasta 7 semanas (Collyer, 1951; Badgley et al., 1955).
Generalmente parecen muy selectivas a la hora de elegir el sitio de la puesta y, habitualmente lo hacen durante la noche.
Los sustratos elegidos son a veces muy específicos (las hembras previamente toquetean vigorosamente con sus palpos la zona
donde depositarán sus huevos), que suelen ser el envés y márgenes de las hojas (ver material ahora citado en Semidalis aleyrodiformis) (Figs. 3, 4), junto/sobre sus nerviaciones, en/bajo cortezas, en la base y parte plana de las acículas, etc., y efectuan las puestas
con huevos bien aislados, en irregulares hileras o en pequeños grupos (generalmente alineados) y adheridos al sustrato sobre
su superficie longitudinal, merced a secreciones de las glándulas de cemento, y quedan adheridos sobre su polo dorsal, no
micropilar, que es el último en salir de las vías genitales femeninas, a veces alternando su forma y adaptándola a la superficie,
y a veces próximos a las colonias de futuras presas (Withycombe, 1922a, 1923, 1925a; Killington, 1936; Narayanan, 1942; Collyer, 1951; Badgley et al., 1955; Muma, 1967, 1971; Yang, 1974; Henry, 1976; Castellari, 1980; New, 1986; Gepp, 1999; Monserrat, 2001; Monserrat et al., 2001; Pantaleoni, 2007, etc.).
Huevos:
Los huevos no son pedunculados, y son elípticos o fusiformes, alargados, subcilíndricos y de extremos redondeados, de unos
0,45-0,55 mm de longitud y unas 2 veces más largos que anchos (Figs. 3, 4, 84). Withycombe, 1922a, 1923, 1924b, 1925a y Killington, 1936 citan 0,4-0,5 mm de longitud en Parasemidalis, Coniopteryx y Conwentzia; Narayanan, 1942 cita 0,5 mm en Coniopteryx; Badgley et al., 1955 citan 0,53 mm en Spiloconis, y Castellari, 1980 cita 0,41-0,44 mm en Coniopteryx, y son algo más aplanados en la superficie donde se adhiere al sustrato donde se depositó el huevo. Son translúcidos o de
tono/ color crema pálido, blanco, amarillo pálido, amarillento, anaranjado, rosáceo, rojizo o pardo al inicio de la puesta
(Badgley et al., 1955; Monserrat et al., 2001). Conforme el tiempo avanza, su color va cambiando según el desarrollo embrionario progresa, y las pardas bandas metaméricas
del embrión primero y los ocelos negros después evidencian su desarrollo (Killington, 1936; Badgley et al., 1955), y el oscurecimiento del huevo los hace menos conspicuos.
|
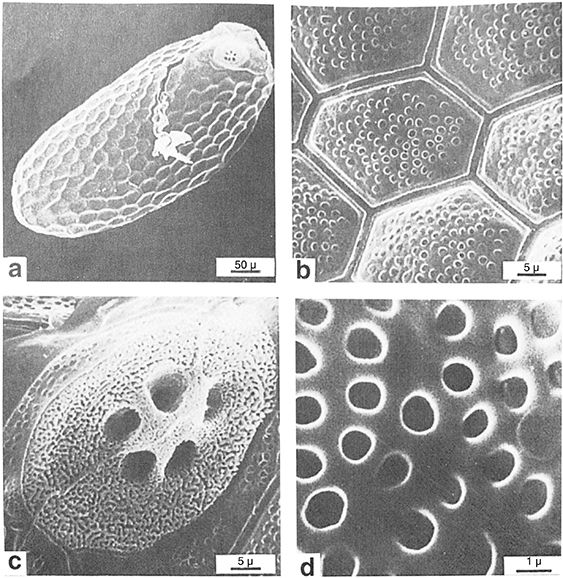 Fig. 25.— Conwentzia psociformis, a: ultraestructura del huevo eclosionado con ovirruptor abandonado, b: aspecto del corion, c: aspecto del micropilo, d: detalle de las estructuras del corion. De Gepp (1990). Fig. 25.— Conwentzia psociformis, a: ultraestructura del huevo eclosionado con ovirruptor abandonado, b: aspecto del corion, c: aspecto del micropilo, d: detalle de las estructuras del corion. De Gepp (1990).
Fig. 25.— Conwentzia psociformis, a: ultrastructure of the hatched egg with abandoned egg-burster, b: chorion appearance, c: micropyle appearance, d: chorion
detailed structures. From Gepp (1990).
|
|
Poseen micropilo aparente (Figs. 3, 4), a veces puntiagudo, en general blanco muy llamativo y con numerosos dispersos aeropilos en él, y poseen frecuentes y pequeñas
impresiones foliculares hexagonales-poligonales sobre el corion (reflejo e impronta de la presión ejercida por las células
foliculares) (Fig. 25) y pueden existir otras menores sobre su superficie que le dan al huevo un aspecto aterciopelado (Withycombe, 1923, 1924b, 1925a; Lacroix, 1924; Killington, 1929, 1936; Stitz, 1931; Narayanan, 1942; Silvestri, 1942; Collyer, 1951; Badgley et al., 1955; Muma, 1967, 1971; Meinander, 1972a; Henry, 1976; Agekyan, 1978; Hinton, 1981; Gepp & Stürzer, 1986; New, 1989; Gepp, 1990; Monserrat, 2001 y material ahora citado en Semidalis aleyrodiformis).
En condiciones experimentales se han citado en diferentes especies puestas de entre 10-20 huevos diarios y puestas de 3-9,
11, 30-40, 22-66, 67-198; 107, 200, 96-266, etc. huevos por hembra (Killington, 1936; Collyer, 1951; Badgley et al., 1955; Monserrat, 2001). Muma (1967) y Stange (1981) citan puestas de 2-5, 9-14 huevos/día en Semidalis vicina, Castellari (1980) cita puestas de 2-8 huevos diarios en Coniopteryx esbenpeterseni, etc., aunque estos datos pueden variar en función de las condiciones, y así Fleschner & Ricker (1953) anotan puestas de 3-9, 22-66, 67-198 huevos en función de la variedad de dietas ofrecidas a las hembras.
Lógicamente el tiempo de desarrollo del huevo varía en función de las condiciones externas, en especial de la temperatura,
oscilando entre 4-5, 6-8, 8-9, 8-12, 8-21, 14, 16 días (Quayle, 1912; Withycombe, 1923; Narayanan, 1942; Badgley et al., 1955; Muma, 1967; Castellari, 1980; Gepp & Stürzer, 1986; Canard & Volkovich, 2001). Withycombe (1923) anota tiempos de desarrollo de 13 días para Conwentzia psociformis, y de 18-21 días en Semidalis aleyrodiformis (6 días en el material ahora citado en Semidalis aleyrodiformis), Withycombe (1924b, 1925a) anota de seis días a dos-tres semanas según temperatura, Killington (1936) anota 1,5-2,5 semanas en los tiempos de desarrollo embrionario en diferentes especies británicas, y Collyer (1951) anota en Conwentzia una media en tiempo de desarrollo del huevo de 5/7/12 días, según los meses del año (VI-VIII/IX/X).
La ruptura del huevo se efectúa mediante una incisión transversal (o longitudinal, según las observaciones) bajo el polo micropilar,
merced al concurso de pequeño ovirruptor embrionario aserrado/denticulado que queda adherido al tegumento (clípeo-labro) embrionario
y al corion abandonado (Fig. 25a) y se realiza merced al concurso de los movimientos del embrión (Withycombe, 1923, 1925a; Killington, 1936; Badgley et al., 1955; Kimmins & Wise, 1962; Muma, 1967; Meinander, 1972a; Castellari, 1980; Gepp & Stürzer, 1986; Gepp, 1990, 1992; Monserrat et al., 2001). El proceso del nacimiento en este tipo de insectos es detalladamente descrito por Killington (1936) y Badgley et al. (1955). Gepp (1990) y Monserrat (2001) recopilan la información existente hasta la fecha sobre el huevo en esta familia.
Estadios larvarios:
Las larvas neonatas permanecen unas horas sobre/junto al huevo antes de iniciar su actividad (Narayanan, 1942; Badgley et al., 1955), siendo éste un periodo especialmente crítico (Canard & Volkovich, 2001). Su desarrollo larvario requiere tres estadios, las sugerencias de cuatro estadios en Heteroconis por Badgley et al. (1955) o en Semidalis por Muma (1967, 1971) no han sido corroboradas y requieren confirmación (personalmente creemos que quizás con este citado “cuarto estadio” se
refieran a la prepupa antes de formarse la pupa real), y de ser ciertas estas observaciones sería, junto a Ithonidae (con
5 estadios), un caso único en esta familia y entre los neurópteros (más de tres fases larvarias). Las larvas neonatas inicialmente
son muy ágiles, veloces y activas, ayudándose con el extremo abdominal para caminar con el que hacen palanca. Se dispersan
desplazándose a gran velocidad buscando su futura fuente de alimentación sobre la vegetación. En este periodo de búsqueda
pueden desplazarse largas distancias y poseen una gran capacidad de sobrevivir sin alimento hasta su muerte (hasta 40 m y
durante más de 10 h cita Fleschner, 1950).
Sus tres fases larvarias son de similares morfologías, aunque obviamente varían en tamaño y quetotaxia, y en general las mayores
diferencias inter-específicas se manifiestan en larvas de tercer estadio. Las larvas son de tipo campodeiforme, generalmente
alargadas, a veces más fusiformes, desnudas (Figs. 5-7) y portadoras de setas de muy diversa morfología (Figs. 27-75) (ver clave de géneros de nuestra fauna). De color blanco, crema, rosáceo o amarillento pálidos, a veces con coloración disruptiva
presentando manchas más oscuras muy contrastadas (Figs. 5-7), en las que se ha mencionado en algunos géneros cierta variabilidad intraespecífica en su coloración (Gepp & Stürzer, 1986; Gepp, 1992).
Aunque en ocasiones se han hallado granos de polen e hifas, esporas y esporangios de hongos en su tracto digestivo (Monserrat, 1998, 2002, 2005b, 2006), son activos depredadores (y excelentes aliados de nuestros intereses) de huevos, ninfas e imagos de pequeños fitófagos
de pequeños tamaños y escasa motilidad, especialmente áfidos, aleyródidos y cóccidos, aunque otros muchos grupos han sido
citados entre sus presas (pseudocóccidos, adélgidos, filoxéridos, diaspídidos, tisanópteros, etc.) (Figs. 5-7), y particularmente ninfas y adultos de ácaros (Oribatida y especialmente Tetranychoidea, Eriophyoidea, Raphignathoidea y
Tarsonemidea: Tetranychidae, Tydeidae, Eriophyidae, Rhyncaphytoptidae, Ereynetidae, Tarsonemidae, Stigmaeidae, etc.), a los
que acceden merced a su pequeño tamaño, y esta adaptación les proporcionan recursos inalcanzables para otros depredadores
de mayor tamaño (Withycombe, 1923, 1924b, 1925a; Killington, 1936; Silvestri, 1942; Collyer, 1951; Kimmins & Wise, 1962; Putman & Herne, 1966; Muma, 1967; Meinander, 1972a; Popov, 1973; Stimmel, 1979; Castellari, 1980; Gepp & Stürzer, 1986; New, 1989; Stelzl, 1989, 1991, 1992; Monserrat, 1996b, 1998, 2005b, 2006; Lo Verde & Monserrat, 1997; Canard, 2001; Gerling et al., 2001; Monserrat et al., 2001; Miller et al., 2004), siendo, también en estadio, excelentes aliados de nuestros intereses contra estos fitófagos (McEwen et al., 2001). Como hemos citado en los imagos, también sus larvas, aunque a veces hallamos especies ampliamente generalistas, en ocasiones
parecen ser muy selectivas, a tenor de la especificidad de plantas soporte sobre las que en particular las hallamos y donde
encuentran sus específicas presas, por lo que, siendo depredadoras, acaban siendo a veces muy selectivas e indirectamente
estrechamente relacionadas con un tipo determinado de planta/s (Withycombe, 1923, 1924b; Killington, 1936; Kimmins & Wise, 1962; Putman & Herne, 1966; Peterson, 1967; Muma, 1967; Stimmel, 1979; Castellari, 1980; New, 1986, 1989; Monserrat & Marín, 1992, 2001; Canard, 2001).
|
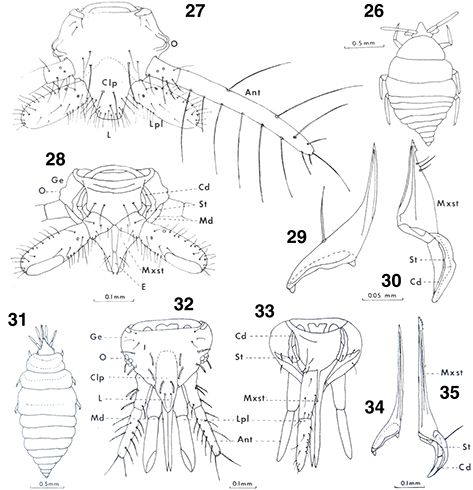 Figs. 26-35.— Morfología de la larva, 26-30: Coniopteryginae, Coniopteryx pygmaea, 26: aspecto general, vista dorsal, 27: cabeza, vista dorsal, 28: ídem, vista ventral, 29: mandíbula, vista ventral, 30:
maxila, vista dorsal, 31-35: Aleuropteryginae, Aleuropteryx loewii, 31: aspecto general, vista dorsal, 32: cabeza, vista dorsal, 33: ídem, vista ventral (omitido un palpo labial), 34: mandíbula,
vista ventral, 35: maxila, vista dorsal (Ant: antena, Cd: cardo, Clp: clípeo, E: epifaringe, Ge: gena, L: labio, Lpl: palpo
labial, Md: mandíbula, Mxst: estilete maxilar, O: ocelos, St: estipe). De Rousset (1966). Figs. 26-35.— Morfología de la larva, 26-30: Coniopteryginae, Coniopteryx pygmaea, 26: aspecto general, vista dorsal, 27: cabeza, vista dorsal, 28: ídem, vista ventral, 29: mandíbula, vista ventral, 30:
maxila, vista dorsal, 31-35: Aleuropteryginae, Aleuropteryx loewii, 31: aspecto general, vista dorsal, 32: cabeza, vista dorsal, 33: ídem, vista ventral (omitido un palpo labial), 34: mandíbula,
vista ventral, 35: maxila, vista dorsal (Ant: antena, Cd: cardo, Clp: clípeo, E: epifaringe, Ge: gena, L: labio, Lpl: palpo
labial, Md: mandíbula, Mxst: estilete maxilar, O: ocelos, St: estipe). De Rousset (1966).
Figs. 26-35.— Larval Morphology, 26-30: Coniopteryginae, Coniopteryx pygmaea, 26: general aspect, dorsal view, 27: head, dorsal view, 28: ditto, ventral view, 29: mandible, ventral view, 30: right maxilla,
dorsal view, 31-35: Aleuropteryginae, Aleuropteryx loewii, 31: general aspect, dorsal view, 32: head, dorsal view, 33: ditto, ventral view (ommited a labial palp), 34: mandible, ventral
view, 35: maxilla, dorsal view (Ant: antenna, Cd: cardo, Clp: clypeus, E: epipharynx, Ge: gena, L: labium, Lpl: labial palpus,
Md: mandible, Mxst: maxillar stylet, O: ocelus, St: stipes). From Rousset (1966).
|
|
Durante la localización de sus recursos se desplazan ayudándose con el extremo del abdomen y mueven lateralmente la cabeza
a uno y otro lado buscando sus presas a las que localizan por contacto directo. Atacan a sus presas sin aparentemente elegir
una posición determinada, especialmente desde abajo (sobre todo huevos y cóccidos acorazados) (Figs. 5-7), y mientras la succionan dirigen los palpos hacia abajo y arquean las antenas hacia atrás (Henry, 1976; Monserrat, 1983a). Debido a sus rectas piezas bucales, su mecanismo de caza difiere de otros neurópteros con mandíbulas-maxilas curvas (Chrysopidae,
Hemerobiidae, Ascalaphidae, Myrmeleontidae, Crocidae, Nemopteridae, etc.), por lo que no pueden “morder” a sus presas, y por
ello prefieren presas sésiles o de escasa moción (Muma, 1967, 1971; Henry, 1976; Stelzl, 1991; Lo Verde & Monserrat, 1997; Canard, 2001), sean huevos, ácaros y homópteros, sean cóccidos, eriófidos o filoxéridos (Figs. 5-7), cuyos fluidos succionan y devoran completamente con suma insistencia (hasta durante 25 minutos por cóccido). Canard (2001) anota una relación de presas observadas en diversas especies por diversos autores. Pueden llegar a ser extremadamente voraces,
habiéndose citado, como ejemplos, Withycombe (1923) quien anota entre 150-300 huevos y ninfas de Phylloxera devoradas a lo largo del desarrollo de Semidalis y Conwentzia; Muma (1967) quien anota hasta casi 3 ácaros por hora devorados por larvas de Semidalis vicina; Quayle (1912, 1913) quien anota en Conwentzia, consumos de hasta 25-53 ácaros de los cítricos al día y consumos de 136-262 ácaros a lo largo de su desarrollo, y hasta 319
ácaros en el tercer estadio de Coniopteryx (Fleschner, 1950), también Karny (1924) menciona esta voracidad en Semidalis pluriramosa. Obviamente el número de presas consumidas va a estar condicionada por el tamaño/sexo del futuro individuo, e influirá en
su longevidad y su potencial reproductor (Fleschner & Ricker, 1953; Canard, 2001). El proceso de la ecdisis es detalladamente descrito por Killington (1936).
Presentan cabeza prognata, pequeña, bien esclerificada (Figs. 26-28, 31, 32, 36-40), y antenas relativamente largas (especialmente en Conwentzia), con abundante y larga setación, formadas por dos segmentos (Figs. 26, 27, 31, 32, 38, 39, 41-43), al igual que los palpos labiales, cuyo segmento distal está fuertemente expandido, y tienen labio muy pronunciado cubriendo
parcial-totalmente las piezas bucales (Figs. 27, 28, 32, 33, 38). Las mandíbulas son rectas, de ápice agudo y no denticulado y de longitud variable según los géneros (Figs. 29, 32-34, 38), y las maxilas son rectas, levemente dentadas en su ápice (Figs. 30, 33, 35) y con canal alimentario entre ambas por donde succionan los líquidos de sus presas (Killington, 1936), ya que el orificio bucal está clausurado por el tegumento tras la eclosión del huevo y vuelve a cerrarse después de cada
ecdisis, por lo que la ingesta debe ser líquida a través de este canal mandíbulas-maxilas (Canard, 2001) y sin duda la secreción de las glándulas salivares ayuda a paralizar a sus presas y predigerir sus fluidos (Rousset, 1966). Carecen de palpo maxilar y el labial es bisegmentado, presentando el más distal una zona fuertemente setosa de carácter
sensorial (Figs. 27, 28, 32, 33 38). Sus ojos poseen cuatro, cinco o seis estematas (Withycombe, 1923, 1925a; Killington, 1936; Rousset, 1966) y frecuentemente la cabeza se encuentra parcialmente embutida en la parte anterior del tórax (Figs. 26, 31). La morfología antenal y de la quetotaxia, así como su disposición sobre la cápsula cefálica son características en cada
género (Figs. 27, 36-41, 80-83), utilizándose en su diagnosis (ver clave de géneros), y se han citado patrones de coloración para distinguir ciertas especies
(Figs. 5-7). Curiosamente Withycombe (1923), Gepp (1992), Gepp & Stürzer (1986) y Sziráki & Flint (2007) encuentran mucha variabilidad intraespecífica en la coloración, bien entre poblaciones o en función de su alimentación,
hecho que no deja de ser sorprendente.
|
 Figs. 36-43.— Morfología y quetotaxia larvaria en diferentes géneros de coniopterígidos: 36: cápsula cefálica de Helicoconis lutea, de Monserrat & Hölzel (1987), 37: cápsula cefálica de Hemisemidalis pallida, de Monserrat & Hölzel (1987), 38: cápsula cefálica de Aleuropteryx loewii, adaptado de Rousset (1966), 39: cápsula cefálica de Semidalis vicina, de Meinander (1974a), 40: cápsula cefálica de Coniopteryx pygmaea, adaptado de Rousset (1966), 41: cápsula cefálica de Conwentzia barretti, de Meinander (1974a), 42: antena de Helicoconis lutea, de Monserrat & Hölzel (1987), 43: antena de Hemisemidalis pallida, de Monserrat & Hölzel (1987). Figs. 36-43.— Morfología y quetotaxia larvaria en diferentes géneros de coniopterígidos: 36: cápsula cefálica de Helicoconis lutea, de Monserrat & Hölzel (1987), 37: cápsula cefálica de Hemisemidalis pallida, de Monserrat & Hölzel (1987), 38: cápsula cefálica de Aleuropteryx loewii, adaptado de Rousset (1966), 39: cápsula cefálica de Semidalis vicina, de Meinander (1974a), 40: cápsula cefálica de Coniopteryx pygmaea, adaptado de Rousset (1966), 41: cápsula cefálica de Conwentzia barretti, de Meinander (1974a), 42: antena de Helicoconis lutea, de Monserrat & Hölzel (1987), 43: antena de Hemisemidalis pallida, de Monserrat & Hölzel (1987).
Figs. 36-43.— Larval Morfology and Chaetotaxy in diferent dusty-wings genera: 36: cephalic capsule of Helicoconis lutea, from Monserrat & Hölzel (1987), 37: cephalic capsule of Hemisemidalis pallida, from Monserrat & Hölzel (1987), 38: cephalic capsule of Aleuropteryx loewii, adapted from Rousset (1966), 39: cephalic capsule of Semidalis vicina, from Meinander (1974a), 40: cephalic capsule of Coniopteryx pygmaea, adapted from Rousset (1966), 41: cephalic capsule of Conwentzia barretti, from Meinander (1974a), 42: antenna of Helicoconis lutea, from Monserrat & Hölzel (1987), 43: antenna of Hemisemidalis pallida, from Monserrat & Hölzel (1987).
|
|
El tórax es sencillo, ancho y transverso, ocupa la mayor parte del conjunto, pero está poco diferenciado del abdomen (Figs. 26, 31, 53, 65, 71). Patas marchadoras bien desarrolladas y de longitud variable (Figs. 26, 31), especialmente largas en Brucheiser o Conwentzia, más cortas en Aleuropteryx o Semidalis, con tibias y un único segmento tarsal no articulado, acabando en dos uñas y un pequeño empodio agudo.
El abdomen consta de diez segmentos de los que los ocho primeros portan estigmas laterales y que, como el tórax, carecen de
especial ornamentación, setación, ni tubérculos, y se encuentran progresivamente adelgazados hacia el extremo caudal (Figs.
26, 31, 53, 65, 71), el cual frecuentemente se utiliza, a modo de palanca sobre el sustrato, para favorecer la locomoción, y también es utilizado
para fijar la larva por este extremo durante la ecdisis y segregar desde los tubos de Malpighi la seda con la que tejer el
capullo pupal (Killington, 1929, 1936; Collyer, 1951) y probablemente también para segregar sustancias defensivas/ofensivas.
El tiempo de desarrollo larvario depende de las condiciones externas, especialmente del alimento y de la temperatura, aunque
hay pocos datos observados sobre los periodos de duración de sus estados larvarios. Se han citado desarrollos de 8-9 días,
de entre 6 días a un par de semanas, de 9-14 días o de 7-20 días en función de la temperatura (Muma, 1967; Castellari, 1980; Stange, 1981). Datos de 41-71 días para completar su desarrollo se han citado para Semidalis aleyrodiformis (Gepp & Stürzer, 1986), de 16-32 días en Spiloconis (Badgley et al., 1955) (con sorprendentes datos sobre cuatro estadios larvarios con duración de 3, 7, 7 y 5-8 días respectivamente), y de 35-43
días en S. vicina (Muma, 1967). En general el primer estadio es breve (3-5 días), el segundo algo mayor (7-10 días) y más largo el tercero (10-19 días).
Withycombe (1923) anota tiempos de 3, 6-7 y 10-11 días, para los tres estadios de Conwentzia psociformis, y de 3-5, 6-7 y 19-20 días en Semidalis aleyrodiformis, Quayle (1912) anota una media de 15 días para el desarrollo en Conwentzia, Collyer (1951) anota también en Conwentzia una media en tiempo de desarrollo larvario de 7/10/16 días según los meses del año (VI-VIII/IX/X), y de entre 16-36 días
desde la emergencia de la larva del huevo a la salida del imago del capullo, igualmente en función de la época del año. También
Narayanan (1942) cita 18-25 días para el completo desarrollo larvario en Coniopteryx, y Canard & Volkovich (2001) aportan una relación de tiempos en el desarrollo de diferentes especies anotados por diferentes autores en diferentes condiciones.
Sobre la morfología de las fases larvarias de las diferentes especies conviene mencionar que, a nivel general, son escasamente
conocidas, desconociéndose en la mayoría/muchas de sus especies (incluso en la fauna europea). Desde que Curtis (1834) describiera
la primera larva, no han sido demasiadas las especies cuyas larvas han sido descritas. Sobre la identificación de las fases
larvarias Monserrat (2001) aporta una clave, a nivel mundial, de las larvas de los géneros que entonces eran conocidos y aporta la información existente
para cada una de las especies cuyas fases juveniles que habían sido descritas, a las que ha de sumarse nuevas descripciones,
siendo especialmente interesante Sziráki & Flint (2007). En nuestra fauna hay algunas referencias sobre los estadios larvarios de alguna de sus especies, que se comentarán en cada
caso (Figs. 26-75), y para el lector interesado, aportamos una sencilla clave general de las larvas ibéricas a nivel de los géneros cuyas larvas
son conocidas. La información sobre la morfología de las fases preimaginales de las especies presentes en la Península Ibérica/Baleares
se anota a lo largo del texto en cada una de ellas, aunque las identificaciones en las descripciones antiguas han de tomarse
con cautela. Para alguna de las especies de las Islas Canarias se puede consultar Monserrat (1978c). Más información sobre estadios preimaginales en Curtis, 1834; Dujardin, 1851; Brauer & Löw, 1857; Löw, 1885; Emerton, 1906; Enderlein, 1906; Quayle, 1912, 1913; Arrow, 1917; Withycombe, 1923, 1925a; Killington, 1936; Narayanan, 1942; Collyer, 1951; Badgley et al., 1955; Schremmer, 1959; Rousset, 1960a, 1966; Kimmins & Wise, 1962; Macleod, 1964; Muma, 1967, 1971; Ward, 1970; Meinander, 1972a, 1974a; Yang, 1974; Greve, 1974; Monserrat, 1978c, 2001; Castellari, 1980; Gepp, 1984, 1986, 1992; Gepp & Stürzer, 1986; Monserrat & Hölzel, 1987; Monserrat et al., 1991; Sziráki & Flint, 2007; Sziráki, 2007b; Zimmermann et al., 2009, etc. Para el lector interesado puede encontrar datos sobre su anatomía en Withycombe (1925a) y Rousset (1956a, 1956b, 1958, 1966, 1968, 1969, 1970).
Pupa:
Apenas hay datos al respecto, pero completado su desarrollo, la larva madura deja de alimentarse, está menos activa e inicia
la pupación. Ésta se realiza mediante la fabricación de un capullo de seda circular o levemente elíptico (Figs. 8, 9, 76), de color blanquecino o amarillento, de aspecto aplanado (parecido al saco ovígero de ciertas arañas tipo Araneida: Heterpodidae-Sparassidae),
de unos 5-8 mm (Badgley et al., 1955 citan 2,5-3 mm de diámetro en Spiloconis). Está formado por dos-tres capas independientes de seda, segregada por los tubos de Malpighi (6 y no 8 como es habitual
en otras familias) y acumulada en un pequeño reservorio, y que sale por el ano, utilizando eficazmente los ágiles últimos
segmentos abdominales para su ejecución y tejido, y las papilas anales (Fig. 52) son utilizadas para este menester (Withycombe, 1924b, 1925a). El proceso de fabricación del capullo es detalladamente descrito por Arrow (1917) y Withycombe (1923). Aunque no siempre suelen elegir zonas protegidas para efectuar la construcción del capullo (p. ej. Conwentzia suele pupar en la superficie de las hojas), suelen preferir zonas más reservadas (Fig. 8), especialmente si se va a producir la hibernación dentro del capullo (si se encuentran en árboles de hoja caduca o marcescente
emigran árboles de hoja perenne o al interior de las cortezas para pupar), sea la base de las acículas, hojas secas y/o recurvadas,
en la base del pedúnculo que sostiene los frutos, bajo cortezas, pies de árboles, etc. (Emerton, 1906; Quayle, 1912; Williams, 1913; Houser, 1914; Withycombe, 1923, 1924b, 1925a; Killington, 1936; Narayanan, 1942; Silvestri, 1942; Collyer, 1951; Badgley et al., 1955; Zimmerman, 1957; Schremmer, 1959; Kimmins & Wise, 1962; Muma, 1967; New, 1968, 1986, 1989; Riek, 1970; Meinander, 1972a; Principi & Canard, 1974; Agekyan, 1978; Ripolles & Melia, 1980; Castellari, 1980; García Marí et al., 1983; Gepp & Stürzer, 1986; Pantaleoni, 2007, etc.). Killington (1936) cita incluso emergencia de imagos de agallas de otros insectos dentro de las cuales habrían realizado su pupación, y esto
lo hemos observado y ahora citamos en algunas de nuestras especies pertenecientes a Aleuropteryx, Coniopteryx, Helicoconis o Hemisemidalis. Según la época del año y especies, la fase de prepupa, fuertemente recurvada sobre sí misma en sus extremos dentro del capullo
(Figs. 9, 77), puede durar 4-5 días desde que se tejió el capullo, antes de pasar a la fase de pupa, o mantenerse en este estadio para
pasar el invierno. Mayoritariamente se ha citado la capacidad de hibernación en fase de prepupa (Withycombe, 1923; Castellari, 1980; New, 1986, 2001a), pero en nuestras latitudes encontramos especies que parecen hacerlo en estado adulto, y muchas otras presentan actividad
a lo largo de todo el año (ver fenologías en cada caso).
|
 Figs. 44-75.— Morfología, pigmentación y quetotaxia larvaria en diferentes especies de coniopterígidos: 44-52: larva de Semidalis candida: 44: aspecto general, vista dorsal, 45: pigmentación cefálica, vista dorsal, 46: quetotaxia cefálica, vista dorsal, 47: extremo
antenal, vista dorsal (setas y órganos placoideos ventrales en punteado), 48: mandíbula, 49: maxila, 50: uña, 51: setas dorsales
del tegumento, 52: extremo abdominal, vista ventral. Adaptado de Monserrat (1978c). 53-60: larva en 3° estadio de Hemisemidalis pallida, 53: aspecto general, ampliadas setas dorsales, 54: región terminal del abdomen, vista dorsal, 55: ídem, vista ventral, 56:
aspecto dorsal de la cápsula cefálica, 57: quetotaxia cefálica dorsal, 58: maxila, vista ventral, 59: mandíbula, vista ventral,
60: antena, vista dorsal. 61-63: larva en 3° estadio de Helicoconis lutea, 61: quetotaxia cefálica dorsal, 62: antena, vista dorsal, 63: aspecto del tegumento abdominal dorsal. Adaptado de Monserrat & Hölzel (1987). 64-69: larva en 3° estadio de Semidalis pluriramosa, 64: aspecto general, vista dorsal, 65: quetotaxia dorsal, porción tegumentaria del pronoto y abdomen ampliada, 66: antena,
vista dorsal, seta del escapo ampliada, 67: pigmentación y quetotaxia de la cápsula cefálica, vista dorsal, setas ampliadas,
68: pigmentación y quetotaxia del extremo abdominal, vista ventral, 69: ídem, vista dorsal. Adaptado de Monserrat et al. (1991). 70-75: larva en 3° estadio de Semidalis pseudouncinata, 70: aspecto general, vista dorsal, 71: quetotaxia dorsal, porción tegumentaria del pronoto ampliada, 72: antena, vista dorsal,
73: pigmentación y quetotaxia de la cápsula cefálica, vista dorsal, seta ampliada, 74: pigmentación y quetotaxia del extremo
abdominal, vista dorsal, 75: ídem, vista ventral. Adaptado de Monserrat et al. (1991). Escalas en mm. Figs. 44-75.— Morfología, pigmentación y quetotaxia larvaria en diferentes especies de coniopterígidos: 44-52: larva de Semidalis candida: 44: aspecto general, vista dorsal, 45: pigmentación cefálica, vista dorsal, 46: quetotaxia cefálica, vista dorsal, 47: extremo
antenal, vista dorsal (setas y órganos placoideos ventrales en punteado), 48: mandíbula, 49: maxila, 50: uña, 51: setas dorsales
del tegumento, 52: extremo abdominal, vista ventral. Adaptado de Monserrat (1978c). 53-60: larva en 3° estadio de Hemisemidalis pallida, 53: aspecto general, ampliadas setas dorsales, 54: región terminal del abdomen, vista dorsal, 55: ídem, vista ventral, 56:
aspecto dorsal de la cápsula cefálica, 57: quetotaxia cefálica dorsal, 58: maxila, vista ventral, 59: mandíbula, vista ventral,
60: antena, vista dorsal. 61-63: larva en 3° estadio de Helicoconis lutea, 61: quetotaxia cefálica dorsal, 62: antena, vista dorsal, 63: aspecto del tegumento abdominal dorsal. Adaptado de Monserrat & Hölzel (1987). 64-69: larva en 3° estadio de Semidalis pluriramosa, 64: aspecto general, vista dorsal, 65: quetotaxia dorsal, porción tegumentaria del pronoto y abdomen ampliada, 66: antena,
vista dorsal, seta del escapo ampliada, 67: pigmentación y quetotaxia de la cápsula cefálica, vista dorsal, setas ampliadas,
68: pigmentación y quetotaxia del extremo abdominal, vista ventral, 69: ídem, vista dorsal. Adaptado de Monserrat et al. (1991). 70-75: larva en 3° estadio de Semidalis pseudouncinata, 70: aspecto general, vista dorsal, 71: quetotaxia dorsal, porción tegumentaria del pronoto ampliada, 72: antena, vista dorsal,
73: pigmentación y quetotaxia de la cápsula cefálica, vista dorsal, seta ampliada, 74: pigmentación y quetotaxia del extremo
abdominal, vista dorsal, 75: ídem, vista ventral. Adaptado de Monserrat et al. (1991). Escalas en mm.
Figs. 44-75.— Larval Morphology, Pigmentation and Chaetotaxy in different coniopterygids species: 44-52: larva of Semidalis candida: 44: general aspect, dorsal view, 45: cephalic pigmentation, dorsal view, 46: cephalic chaetotaxy, dorsal view, 47: antennal
apex, dorsal view (setae and ventral placoid organs puncted), 48: mandible, 49: maxilla, 50: claw, 51: dorsal tegument setae,
52: abdominal apex, ventral view. Adapted from Monserrat (1978c). 53-60: 3rd instar larva of Hemisemidalis pallida, 53: general aspect, magnified dorsal setae, 54: abdomimal apex, dorsal view, 55: ditto, ventral view, 56: dorsal aspect
of cephalic capsule, 57: dorsal cephalic chaetotaxy, 58: maxilla, ventral view, 59: mandible, ventral view, 60: antenna, dorsal
view. 61-63: 3rd instar larva of Helicoconis lutea, 61: dorsal cephalic chaetotaxy, 62: antenna, dorsal view, 63: dorsal abdominal tegument aspect. Adapted from Monserrat &
Hölzel (1987). 64-69: 3rd instar larva of Semidalis pluriramosa, 64: general aspect, dorsal view, 65: dorsal chaetotaxy, pronotus and abdominal tegument magnified, 66: antenna, dorsal view,
scapus seta magnified, 67: pigmentation and chaetotaxy of cephalic capsule, dorsal view, setae magnified, 68: pigmentation
and chaetotaxy of abdominal tip, ventral view, 69: ditto, dorsal view. Adapted from Monserrat et al. (1991). 70-75: 3rd instar larva of Semidalis pseudouncinata, 70: general aspect, dorsal view, 71: dorsal chaetotaxy, pronotus tegumentary portion magnified, 72: antenna, dorsal view,
73: cephalic capsule pigmentation and chaetotaxy, dorsal view, seta magnified, 74: tip abdominal pigmentation and chaetotaxy,
dorsal view, 75: ditto, ventral view. Adapted from Monserrat et al. (1991). Scales in mm.
|
|
La pupa es de tipo dectica y exarata, con mandíbulas simétricas, planas, portadoras de una pequeña muesca o hendiduran el
margen interno, y las antenas son libres y dispuestas alrededor de la cabeza hacia las pterotecas (Figs. 78, 79), hecho habitual en los Neuropterida (Emerton, 1906; Withycombe, 1923, 1925a; Killington, 1936; Greve, 1974). Datos sobre pupas en diferentes especies en Emerton (1906), Withycombe (1924b), Stitz (1931), Killington (1936), Badgley et al. (1955), Greve (1974), Gepp & Stürzer (1986), Monserrat (2001), etc.
El periodo pupal es relativamente corto, y si no hay hibernación en este estado, la emergencia del imago, generalmente al
atardecer, acontece a los 8-10 días en meses cálidos y hasta los 30 días en meses más frescos, y de 10-12/11-20 días en condiciones
de laboratorio y tras 5-8, 11-12, 15 días, según otras observaciones. A diferencia de lo acontecido en otras familias (Mantispidae,
Berothidae, Hemerobiidae, Sisyridae u Osmylidae), la salida del capullo no se ve favorecida por la posesión de pares de dentículos
curvos y opuestos sobre la región tergal de algunos segmentos del abdomen pupal que actúan como anclaje para liberarse de
la exuvia pupal (Killington, 1936), que no existen en esta familia (Figs. 78, 79), aunque la propia exuvia pupal abandonada actúa de anclaje (Badgley et al., 1955). Para liberarse del capullo de seda la pupa la corta con sus mandíbulas un orificio circular-semicircular (Withycombe, 1923; Badgley et al., 1955) y, tras retirarse un trecho, deja abandonada en él su exuvia pupal (pocas veces se ha observado salir del capullo la pupa
entera: Badgley et al., 1955), y tras liberarse de ella, queda libre el imago, que suele trepar por una superficie vertical, y tras expeler varias gotitas
de fluido viscoso (restos del contenido del tubo digestivo que no ha podido ser eliminado por el ano con anterioridad, debido
a la falta de conexión entre el mesodeo y proctodeo) y no merced a una única estructura sólida (meconio) que caracteriza a
los estados juveniles de las otras familias de neurópteros (Withycombe, 1925a; Killington, 1936; Canard & Volkovich, 2001; Canard, 2001), comienza el endurecimiento tegumentario y la expansión alar mientras frota sus alas en expansión con las patas posteriores
(Withycombe, 1924b). Posteriormente se inicia, entre media hora y 48 horas, la secreción y el recubrimiento activo de la cera que empiezan a
segregar sus glándulas tegumentarias, en poco más una hora está listo para volar. El proceso de emergencia del imago es detalladamente
descrito por Withycombe (1923, 1925a), Killington (1936) y Badgley et al. (1955). Información general sobre la anatomía, morfología y biología de las especies de esta familia (huevos, larvas, pupas e imagos)
puede recabarse en la bibliografía citada adjunta.
|
 Figs. 76-84.— Aspectos de las fases juveniles de coniopterígidos: 76-78: Semidalis vicina: 76: capullo, vista dorsal y sección, 77: prepupa, 78: pupa, de Emerton (1906), 79: pupa de Helicoconis lutea, de Greve (1974), 80-83: tipos de setas en diferentes géneros: 80: Heteroconis, de Badgley et al. (1955), 81, 82: Hemisemidalis, de Monserrat & Hölzel (1987), 83: Semidalis, de Monserrat et al. (1991), 84: Aleuropteryx iberica, huevo, de Monserrat (1977b). Escalas para figs. 79 y 84. Figs. 76-84.— Aspectos de las fases juveniles de coniopterígidos: 76-78: Semidalis vicina: 76: capullo, vista dorsal y sección, 77: prepupa, 78: pupa, de Emerton (1906), 79: pupa de Helicoconis lutea, de Greve (1974), 80-83: tipos de setas en diferentes géneros: 80: Heteroconis, de Badgley et al. (1955), 81, 82: Hemisemidalis, de Monserrat & Hölzel (1987), 83: Semidalis, de Monserrat et al. (1991), 84: Aleuropteryx iberica, huevo, de Monserrat (1977b). Escalas para figs. 79 y 84.
Figs. 76-84.— Aspects of Juvenile Stages in dusty-wings: 76-78: Semidalis vicina: 76: cocoon, dorsal view and section, 77: prepupa, 78: pupa, from Emerton (1906), 79: pupa of Helicoconis lutea, from Greve (1974), 80-83: kinds of setae in diferent genera: 80: Heteroconis, from Badgley et al. (1955), 81, 82: Hemisemidalis, from Monserrat & Hölzel (1987), 83: Semidalis, from Monserrat et al. (1991), 84: Aleuropteryx iberica, egg, from Monserrat (1977b). Scales for figs. 79 and 84.
|
|
Fenología:
Mayoritariamente en nuestras latitudes las poblaciones se desarrollan durante los meses de primavera y verano, sucediéndose
varias (2, 3, 4) generaciones en función de la temperatura, fotoperiodo, etc., y de los recursos del medio, y la hibernación
suele producirse en fase de prepupa, más raramente en pupa, y ocasionalmente en cualquier fase desde huevo a imago (Killington,
1936; Marín & Monserrat, 1987; New, 1989; Marín, 1994). En zonas de la península litorales y más meridionales las poblaciones de ciertas especies se desarrollan y suceden a lo
largo de todo el año, incluso ya pasado el otoño en los meses de invierno. La mayoría de nuestras especies parecen tener ciclos
bivoltinos, pero incluso más (Killington, 1936; Castellari, 1980; Pantaleoni, 1983, 1990, 2007; Gepp & Stürzer, 1986; New, 2001b; Penny et al., 2007), aunque en ciertas condiciones también ciclos monovoltinos están citados en especies adaptadas a medios, latitudes o altitudes
más rigurosos (p. ej.: Helicoconis lutea), aunque en algunos casos esto puede variar al adaptarse a la latitud y la altitud de las poblaciones. En nuestras latitudes
se han citado hasta 4 generaciones anuales en especies como Coniopteryx haematica, y probablemente esto ocurra en otras especies de Coniopteryx, Semidalis o Conwentzia (Marín & Monserrat, 1987), y así lo demostramos con el material ahora estudiado de algunas especies (Conwentzia psociformis, Parasemidalis fuscipennis, Coniopteryx haematica, Semidalis aleyrodiformis) que, en zonas litorales y de influencia mediterránea, muestran una sucesión de generaciones a lo largo de todo el año. En
zonas tropicales y/o subtropicales las generaciones se suceden sin ninguna diapausa y, en cualquier caso, la relativamente
más larga longevidad de las hembras (Collyer, 1951; Castellari, 1980) y de su periodo reproductor hacen dificultoso calibrar el número de generaciones reales en las poblaciones de la mayoría
de nuestras especies, dado que se solapan las poblaciones derivadas del periodo reproductor de una misma hembra. Killington (1936: 148) anota las fenologías de las especies del Reino Unido.
Relaciones interespecíficas:
Para finalizar este apartado de generalidades citemos que, obviamente, los coniopterígidos también son presas (como larvas
o imagos) de otros artrópodos depredadores, principalmente ácaros, pequeñas arañas, diplópodos, melíridos, antocóridos, larvas
de coccinélidos y sírfidos, hormigas, e incluso larvas e imagos de crisopas, etc., así como de otros animales predadores de
mayor tamaño como anfibios, reptiles, aves y murciélagos (Laidlaw, 1923; Withycombe, 1923, 1924b; Lacroix, 1924; Killington, 1936; New, 1967, 1975, 1986, 1989; Miller et al., 2004, etc.).
Por último citar que diversos tipos de parásitos e hiperparásitos han sido citados en esta familia, especialmente sobre sus
huevos, sus larvas o emergiendo de sus capullos, incluso parasitando imagos, especialmente himenópteros parásitos, parasitoides
e hiperparásitos: Hymenoptera, Apocrita, Chalcidoidea: Aphelinidae (Aphelinus), Eulophidae (Tetrastichus); Ichneumonoidea: Ichneumonidae (Calliceras), Braconidae (Rhizarcha); Ceraphronoidea: Ceraphronidae (Ceraphron, Lygocerus, Aphanogmus, Dendrocerus, etc.), a veces solo citados como Ceraphronidea, Chalcididae, Megaspilidae, Encyrtidae o Eulophidae (Arrow, 1917; Lacroix, 1924; Withycombe, 1923, 1924b; Killington, 1932, 1936; Eglin, 1940; Silvestri, 1942; Schremmer, 1959; Viggiani, 1967; New, 1967, 1989; Tjeder, 1969; Askew, 1971; Dessart, 1978, 1998; Pantaleoni, 1984; Sinacori et al., 1992, etc.), alcanzando porcentajes de hasta el 40-50 % (Castellari, 1980; New, 1986). También ciertas gregarinas y ciertas bacterias se han citado en larvas, pupas e imagos (Withycombe, 1923).
HISTORIAL PALEONTOLÓGICO
Sobre su historial paleontológico, y a diferencia de otras familias de neurópteros, este grupo por su especial morfología
y pequeño tamaño, es de mayor dificultad a la hora de dejar su impronta pretérita (mayoritariamente en ámbar) y asignarlos
en ejemplares fósiles (Tillyard, 1932; Jepson & Penney, 2007), aunque no son escasos los registros de coniopterígidos fósiles (Hagen, 1856; Handlirsch, 1906-1908, 1909; Enderlein, 1909, 1910a, 1930; Carpenter, 1930, 1992; Killington, 1936; Meinander, 1972a, 1975a, 1979a, 1990; Schlüter, 1986; Nel, 1991; Ohm, 1995; Ansorge, 1996; Dobosz & Krzeminski, 2000; Azar et al., 2000; Engel, 2002, 2004, 2010; Grimaldi & Engel, 2005; Nel et al., 2005; Engel & Grimaldi, 2007, 2008; McKellar et al., 2008; Kupryjanowicz & Makarkin, 2008; Pérez-de la Fuente, 2012; Perrichot et al., 2014, etc.), y parecen haberse diferenciado del resto de los neurópteros (quizás excepción hecha de Ithonidae) desde hace mucho
tiempo (Grimaldi & Engel, 2005). En relación con ello, Meinander (1992) opinaba que desde el Jurásico-Cretácico (Aleuropteryginae) y algo después Coniopteryginae, y Winterton et al. (2010) sostienen su diferenciación como grupo hermano del resto de los neurópteros del que divergió hace c. 294 ma durante finales
del Carbonífero-principios del Pérmico, diversificándose las dos principales subfamilias actuales (Coniopteryginae, Aleuropteryginae)
durante el Triásico.
En cualquier caso, los registros fósiles ofrecen en general aspectos muy similares a los actuales, hecho que refleja una marcada
parsimonia. Sus registros parecen bien diferenciados desde el Mesozoico. El primer registro asignable a esta familia corresponde
a †Juraconiopteryx zherichini del Jurásico Superior de Kazakstán, si bien también se ha relacionado con los homópteros. De posición más incuestionable
citemos †Libanosemidalis hammanaensis y †Alboconis cretacica del Cretácico Inferior del Líbano, †Glaesoconis cretica del ámbar cretácico siberiano, †Glaesoconis baliopteryx y †Phthanoconis burmitica del ámbar cretácico de Myanmar, †Glaesoconis fadiacra del ámbar cretácico libanés (transferida al género †Libanoconis), †Garnaconis dupeorum del ámbar cretácico francés, y †Glaesoconis nearctica, †Apoglaesoconis ackermani, †Apoglaesoconis swolenskyi, †Apoglaesoconis luzii, †Apoglaesoconis cherylae del ámbar Turoniano de New Jersey, y también ejemplares de †Glaesoconis han sido encontrados en el ámbar cretácico de El Soplao (Cantabria), y otros de ubicación discutida dentro de esta familia
(†Archiconiopteryx liasina, inicialmente descrita dentro del género †Archipsylla y recientemente transferida a Homoptera: †Archiconiopterygidae) del Jurásico europeo, existiendo otros registros posteriores del Cenozoico donde aparecen más diversificados y son aparentemente
más frecuentes. Tal es el caso de †Archiconiocompsa prisca del Eoceno-Oligoceno en ámbar báltico y de Ucrania, y más recientes otros géneros monoespecíficos (†Archiconis, †Heminiphetia) del ámbar báltico del Oligoceno-Eoceno y †Pararchiconis quievreuxi del Oligoceno francés y †Gallosemidalis eocenica del Eoceno francés, †Spiloconis eominuta del Eoceno indio, †Hemisemidalis sharovi y †Hemisemidalis kulickae del Eoceno báltico. En cualquier caso, mayoría de los datos son relativamente recientes (Oligoceno-Mioceno), en ámbar báltico,
dominicano y subfósiles en copal africano (Togo, Madagascar), donde hallamos géneros actuales (Hemisemidalis, Coniopteryx, Semidalis, Neoconis, Spiloconis). Más información en Hagen, 1856; Handlirsch, 1906, 1906-1908, 1909; Enderlein, 1906, 1909, 1910a, 1930; Meunier, 1910a, 1910b, 1910c; Krüger, 1917; Carpenter, 1930; Meinander, 1972a, 1975a, 1979a, 1981, 1998c; Larsson, 1978; Whalley, 1980; Schlüter, 1986; New, 1989; 2001b; Nel, 1991; Carpenter, 1992; Ansorge, 1996; Dobosz & Krzeminski, 2000; Grimaldi, 2000; Azar et al., 2000; Grimaldi et al., 2002, 2013; Engel, 2002, 2004, 2010; Nel et al., 2005; Grimaldi & Engel, 2005; Jepson & Penney, 2007; McKellar et al., 2008; Kupryjanowicz & Makarkin, 2008; Winterton et al., 2010; Najarro et al., 2010; Pérez-de la Fuente, 2012; Grimaldi et al., 2013; Perrichot et al., 2014, etc., y especialmente destacamos Engel (2004) y Engel & Grimaldi (2007, 2008) quienes listan las especies entonces conocidas.
BREVE HISTORIAL SOBRE SU TAXONOMÍA Y SU PROGRESIVO CONOCIMIENTO
El concepto linneano de “neuroptera” incluía numerosos grupos de insectos, tanto heterometábolos como holometábolos (Odonata,
Plecoptera, Ephemeroptera, Isoptera, Psocoptera, Embioptera, Mecoptera, Trichoptera), incluyendo los tres órdenes que hoy
constituyen los Neuropteroidea (Handlirsch, 1906-1908): Neuroptera, Raphidioptera y Megaloptera, y que desde Linnaeus (1758) se han ido desglosando en diferentes órdenes y estos
en las familias actualmente reconocidas (aún algunas: Osmylidae, Sisyridae, Nevrorthidae, Crocidae, etc., tardarían en constituirse),
conforme se realizaban los primeros intentos en organizar su taxonomía y su sistemática (Poda, 1761; Scopoli, 1763; Pallas, 1772; Fabricius, 1775, 1777, 1778, 1781, 1793, 1798; De Geer, 1778, 1783; Schrank von Paula, 1781; Rossi, 1790; Olivier, 1792; Rambur, 1842; Brauer, 1852, 1855a, 1855b; Brauer & Löw, 1857; Packard, 1883a, 1883b, etc.). Ver un buen análisis de esta evolución en Krüger (1915), y para información sobre el concepto linneano de los Neuropteroidea y su evolución se recomienda Bodenheimer (1929), Tjeder (1952), New (1989) y Pantaleoni (2010).
Obviamente el historial de la familia Coniopterygidae, ha ido paralelo a la propia evolución de su concepto, del de la Entomología,
del de la Taxonomía y la Nomenclatura modernas, y de la Neuropterología en particular, conforme el primevo concepto de “neuróptero”
se iba desglosando en algunos taxa supragenéricos y en familias distintas. Circunscribiéndonos a la familia que nos ocupa,
el concepto de los “actuales” coniopterígidos estaba mezclado dentro de varios órdenes/familias actuales y, dada la particular
morfología de estos insectos, fue especialmente dificultoso adjudicarlo dentro de una categoría taxonómica o asociarlos con
otros géneros entonces conocidos.
Considerados inicialmente como pertenecientes o asociados a Hemerobiidae-Hemerobiinae (Westwood, 1834; Wesmael, 1836a, 1836b, 1841; Walker, 1853; Brauer & Löw, 1857), también estuvieron incluidos o junto a Embioptera, Isoptera, Psocoptera, Ephemeroptera y Odonata. Las primeras especies
se asociaron al entonces ambiguo y diverso género Hemerobius Linnaeus, 1758 (las primeras especies fueron descritas dentro de este género: Hemerobius parvulus Müller, 1764, Hemerobius pulchellus Block, 1799), a las que siguieron otras posteriormente asociadas a esta familia (Meinander, 1972a), aunque algunas otras dentro de, o vinculadas a, géneros hoy día taxonómicamente muy alejados (Trichoptera, Homoptera, Psocoptera,
Megaloptera, etc.): Phryganea Fabricius, 1795 (Phryganea alba Fabricius, 1789), Salmacis Zetterstedt, 1840 (Salmacis hepiolina Zetterstedt, 1840 nomen nudum), Semblis Fabricius, 1775 (Semblis farinosa Rossi, 1794) y a otros géneros ya más cercanos: Sciodus Zetterstedt, 1840, Malacomyza Wesmael, 1836, Aleuronia, Fitch, 1854, Coniortes Westwood, 1834, etc.
Poco a poco van apareciendo primeras clasificaciones y monografías que van centrando la posición sistemática de los neurópteros
y ampliando su conocimiento, y poco a poco el concepto de la familia Coniopterygidae iba adquiriendo entidad (Leach, 1815; Cuvier, 1817; Billberg, 1820; Curtis, 1834; Westwood, 1834, 1840; Newman, 1835, 1853; Wesmael, 1836a, 1936b, 1841; Pictet, 1836; Burmeister, 1839; Rambur, 1842; Hagen, 1851, 1852, 1854, 1856, 1858a, 1858b, 1859a, 1859b, 1861, 1866, 1873; Brauer, 1852, 1855a, 1855b, 1868, 1876, 1900; Walker, 1853; Fitch, 1854, 1856; Brauer & Löw, 1857; Wallengren, 1863, 1871; McLachlan, 1867, 1868, 1870a, 1870b, 1880, 1882a; Löw, 1885; Rostock & Kolbe, 1888; Albarda, 1889; Banks, 1892, 1899; Reuter, 1894; Vine, 1895; Perkins, 1899, etc.), que entroncan con los autores del pasado siglo. La familia Coniopterygidae fue definida por Burmeister (1839: 771)
con el género Coniopteryx como género tipo, y dos especies entonces a ella/él asignadas: C. tineiformis y C. psociformis (con la misma intención, poco después Newman, 1853: 201 la definiría y llamaría Coniopterycidae) y paralelamente Hagen (1866)
también elevaría a nivel de familia muchos taxa anteriormente incluidos en la por entonces heterogénea familia Hemerobiidae
(Myrmeleontides, Ascalaphides, Nemopterides, Mantispides, Hemerobiides, Chrysopides, y Coniopterygides). Para los estudios
iniciales sobre esta familia se recomienda Aspöck & Aspöck (2003) y Zimmermann (2005b).
Las obras de Enderlein (1905a, 1905b, 1905c, 1906, 1907a, 1907b, 1907c, 1908, 1909, 1910a, 1910b, 1914, 1929, 1930) sentaron las bases generales de la sistemática y la nomenclatura morfológica de esta familia que ha sido habitualmente aceptada,
aunque es raro no encontrar muy diversos autores que han modificado o propuesto su propia nomenclatura, hecho que complica
la terminología a emplear. Tuxen (1970) utiliza más de 4000 términos en las estructuras genitales de los insectos, de los que unos 200 son aplicados al orden que
tratamos, y Tjeder (1954, 1956, 1970) y Acker (1960) tratan de uniformizar la terminología genital de este orden, aunque como decimos pocos autores no han modificado la existente
y han propuesto sucesivamente la suya propia. Nosotros mantenemos la utilizada en su revisión mundial de esta familia por
Meinander (1972a).
A partir de la excepcional obra de este entomólogo alemán Günther Enderlein [1872-1968], la taxonomía de esta familia se vió
reforzada por la utilización de caracteres de genitalia que definieran las especies, y no sólo en base a variables caracteres,
mayoritariamente de venación, que hasta entonces (y posteriormente) habían venido siendo utilizados para diferenciar los diferentes
taxa. Sin embargo sorprendentemente muchos autores (lamentablemente en nuestro caso L. Navás) mayoritariamente no la tuvieron
en cuenta, y siguieron utilizando elementos de morfología externa muy variables y poco recomendables, contribuyendo a la confusión
en la taxonomía y faunística de muchas especies (ver más adelante reseña a las citas de Navás y otras citas antiguas en nuestras
especies). Para la evolución taxonómica y sistemática de la familia Coniopterygidae se recomienda Meinander (1972a).
Desde las primeras monografías (Enderlein, 1906, 1908) numerosas especies y géneros de esta familia han venido describiéndose, habiendo sido recogidas en la más reciente revisión
(Meinander, 1972a). El catálogo de las especies del mundo fue aportado por Meinander (1990), actualizado por Sziráki (2011), y la revisión de las especies europeas y la relación de especies de la Región Paleártica occidental han sido recogidas por
H. Aspöck et al. (1980, 2001), y Sziráki (2004) aporta una clave que incluye las especies de Eurasia y norte de África, y Sziráki (2011) mundiales, anotando en ambos casos las referencias recientemente aparecidas. Para el lector interesado, más adelante anotamos
los trabajos más relevantes con información sobre la taxonomía y la sistemática de esta familia. Para los interesados en conocer
información sobre los géneros de esta familia puede consultarse Oswald & Penny (1991), y sobre todos sus taxa y la bibliografía de esta familia se recomienda Oswald (2013a, 2013b).
Sin duda, las faunas Paleártica, Neártica y Australiana son las mejor conocidas sobre este particular, si bien el límite real
de distribución de muchas especies es, con frecuencia, impreciso o incompleto y, en ocasiones, los datos sobre su biología
son escasos y el nivel de conocimiento de las faunas de muchos países, incluso en zonas relativamente bien estudiadas como
Europa y Estados Unidos y recientemente China, es aún poco satisfactorio. Este hecho es aún más evidente en faunas de otros
continentes, y nuevas obras han sido afrontadas, y desde el último tercio del pasado siglo al presente nuevos trabajos van
aportando nuevos datos que van completando las lagunas existentes sobre su taxonomía, distribución, faunística, biología y
catalogación, siendo ingente el número de publicaciones que han aparecido sobre faunas ajenas a lo que pretende esta contribución.
Al margen de las especies fósiles mencionadas, la familia está dividida en tres subfamilias de especies actuales: La aberrante
y curiosa Brucheiserinae, de distribución neotropical, habitante de zonas desérticas del norte de Argentina y Chile, inicialmente
considerada con categoría de familia e inicialmente incluida en Megaloptera (Navás, 1927a), y Sziráki (2007a) encuentra nuevas homologías entre ambos taxa, con 4 especies (dos de ellas podrían estar basadas en diferencias entre los
sexos de una misma especie) y 2 géneros de morfologías muy diferentes y con venación alar muy particular: aberrante, con vena
costal y subcostal fusionadas, y ambas a la vena radial salvo en su base alar, y venación marcadamente reticulada en Brucheiser, o de venación subparalela, con numerosas venillas transversales, especialmente en las alas posteriores y dos basales en
el campo costal en Flintoconis (Navás, 1927a; Riek, 1975; New, 1987, 2001b; Meinander, 1972a, 1990; Sziráki, 2007a), que desde nuestro punto de vista hace poco sostenible la inclusión de ambos géneros en una misma subfamilia (a pesar de
los argumentos de Sziráki, 2007a). Zimmermann et al. (2009) consideran a Brucheiserinae más próxima a Coniopteryginae que a Aleuropteryginae. Al margen de ésta, tenemos las dos subfamilias
más extensas: Coniopteryginae y Aleuropteryginae, subdivididas a su vez en varias tribus y varios géneros (unos 22 géneros)
y alguno de ellos en subgéneros (Enderlein, 1905a, 1905b, 1905c, 1906, 1908; Meinander, 1972a, 1990, 1992; New, 2001b; Grimaldi et al., 2013), con venación de tipo longitudinal y bastante reducida (Figs. 23, 25) y característica en su disposición, importante para la diferenciación de sus taxa supraespecíficos (ver clave de subfamilias
y géneros), así como, entre otras, diferencias en la morfología de la galea y de las larvas, y que con distribución cosmopolita
representan el grueso de la familia. Meinander (1972a, 1979a) ofrece un excelente y amplio comentario sobre sus diferencias, homologías, relación y posible filogenia y biogeografía de
estas dos subfamilias y entre los géneros entonces conocidos, y Meinander (1981) sobre el género Coniopteryx, actualizando la lista de sus especies. Todas nuestras especies son representantes de estas dos últimas subfamilias (Aleuropteryginae,
Coniopteryginae).
Por si fuera de interés al lector, entre los autores y obras destacables publicadas desde el pasado siglo, ya hemos mencionado
las primeras y excelentes monografía de esta familia Enderlein (1906, 1908), y posteriores obras merecen mencionarse Dziędzielewicz (1905, 1907), Banks (1906, 1913), Perkins (1910), Nakahara (1913), Krüger (1915), Bagnall (1915a, 1915b), Roepke (1916, 1917), Esben-Petersen (1918, 1928), Tillyard (1919, 1926, 1932), Withycombe (1922a, 1922b, 1924a, 1924b, 1925a, 1925b), Karny (1923, 1924), Lacroix (1924), Stitz (1927, 1928, 1931), Carpenter & Lestage (1928), Lackschewitz (1929), Smith (1934), Kimmins (1934, 1935, 1950, 1952, 1953, 1960); Klingstedt (1936), Killington (1936, 1937a, 1937b), Yang (1951, 1964, 1974), Fraser (1952, 1955, 1957, 1959), Carpenter (1955), Zelený (1961a, 1961b, 1964), Ward (1961), Kimmins & Wise (1962), Kuwayama (1962), H. Aspöck (1963a, 1963b, 1964, 1992), Ohm (1963, 1965, 1968, 1973a, 1984, 1986, 1995), Kis (1964a, 1964b, 1965a, 1965b, 1967, 1969), Aspöck & Aspöck (1964, 1965a, 1965b, 1969, 1999, 2003, 2007, 2008, 2010, 2014), Alayo (1968), Gepp (1969), Kis et al. (1970), Adams (1973), Greve (1974), Riek (1975), Johnson (1976, 1977, 1978, 1980a, 1980b, 1981), Ghosh & Sen (1977), Rausch & Aspöck (1977, 1978a, 1978b, 1978c), Rausch et al. (1978a, 1978b, 1981), H. Aspöck et al. (1980, 2001), Panfilov (1980), Monserrat (1981a, 1981b, 1982c, 1983a, 1984f, 1984g, 1989a, 1989b, 1995, 1996b, 1998, 2002, 2005b, 2006, 2010, 2011), Eglin (1982a, 1982b), González Olazo (1984a, 1984b, 1987), Mansell (1986); New (1987, 1988, 1990, 1998, 2001b), Zakharenko (1987, 1988, 1989), Frazer (1988), New & Sudarman (1988); Monserrat & Díaz-Aranda (1988b); Sziráki (1989, 1990, 1991, 1992a, 1992b, 1992c, 1992d, 1992e, 1994a, 1994b, 1996, 1997, 1998a, 1998b, 1999, 2001, 2002a, 2002b, 2004, 2005, 2007a, 2007b, 2009, 2010a, 2010b, 2010c, 2011, 2015), Makarkin (1990), Monserrat et al. (1991), Oswald & Penny (1991), Plant (1991), U. Aspöck (1992, 1995, 2002, 2003), Yang & Liu (1993a, 1993b, 1994, 1999), Liu & Yang (1993, 1997, 1998, 2001, 2002, 2006), Günther (1993), Duelli (1994b), Aspöck & Mansell (1994), Monserrat & Bayo (1995), Liu (1995, 2003), Aspöck & Hölzel (1996), Sziráki & Greve (1996, 2000), Stange & Wang (1998), U. Aspöck et al. (2001, 2003, 2012, 2015), Liu et al. (2003a, 2003b, 2004a, 2004b), Zimmermann (2005a, 2005b), Sziráki & Harten (2006), Sziráki & Winterton (2012), Sziráki & Penny (2012), Sziráki & Mirmoayedi (2012), Grimaldi et al. (2013), Vidlička (2014, 2015), Lock (2015), Martins & Amorim (2016), etc., que han aportado interesantes contribuciones, y algunos nuevos elementos vienen publicándose recientemente.
Además de la mencionada obra de Enderlein, y al margen de todas estas contribuciones citadas, es de destacar la obra extensa
del finlandés Martin Meinander [1940-2004] (Meinander, 1962a, 1962b, 1963, 1965a, 1965b, 1969a, 1969b, 1971, 1972a, 1972b, 1972c, 1973, 1974a, 1974b, 1974c, 1974d, 1975a, 1975b, 1975c, 1975d, 1976, 1977, 1979a, 1979b, 1980, 1981, 1982, 1983a, 1983b, 1986, 1990, 1992, 1995, 1996, 1998a, 1998b, 1998c; Meinander & Penny, 1982).
Tras toda esta información, mencionemos que, a diferencia de la mayoría de las grandes familias de neurópteros, esta familia
posee un excelente nivel de conocimiento global en lo que se refiere a su taxonomía, a pesar de no ser especialmente abundante
en colecciones. Desde la obra de Enderlein (1906, 1908), quien recoge 9 géneros y 39 especies, entonces considerados, a nivel general la familia fue posteriormente revisada por
Meinander (1972a) quien recoge 231 especies actuales válidas, y aunque numerosas sinonimias habían venido siendo propuestas, en su posterior
obra (Meinander, 1990) esta cifra se eleva a 423 el número de especies actuales válidas. La cifra actual alcanza las 550 especies (Sziráki, 2004) y no para de crecer (Sziráki, 2011). Como hemos indicado, las especies europeas fueron mayoritariamente revisadas por Aspöck et al. (1980).
En relación con la taxonomía de esta familia, creemos que es la ocasión de anotar unas últimas reflexiones personales que
desde hace tiempo consideramos necesarias comentar. Inevitablemente la opinión de cada taxónomo no deja de ser subjetiva y
variable según su formación, su criterio y su intencionalidad, y estos factores inciden en la Taxonomía en general y en la
de los neurópteros y en la de los coniopterígidos que ahora tratamos en particular. Es sobradamente conocida la extendida
práctica en la “ávida” descripción de nuevos taxa como supuesto método de obtener un mayor “prestigio científico” y con ello
ser “mejor o peor taxónomo” en función del número de nuevas especies descritas por cada cual, y también es sobradamente conocida
la histórica presión que se ejerce en la descripción de nuevos taxa con la aparente intención de “rentabilizar” expediciones
o campañas de instituciones y museos. En base a estos supuestos, varios autores parecen haber abandonado la línea marcada
por Enderlein (1906) y Meinander (1972a) (ver sus artículos en la bibliografía), y la taxonomía de esta familia se ha visto en parte alterada/magnificada con la
descripción de numerosas nuevas especies en base a mínimos caracteres externos o genitales que no son más que elementos propios
de la lógica variabilidad de las poblaciones, de los individuos o de su actividad vital o sexual, sin tener en cuenta la esperable
variabilidad dentro de las poblaciones (especialmente frecuente a nivel intraespecífico en muchos géneros de esta familia),
ni el grado de madurez de los individuos (la esclerificación de ciertas estructuras empleadas con frecuencia en la diagnosis
de nuevas especies cambia o puede cambiar en los individuos en función de su edad y su actividad sexual previa), y hace sospechosa
y dudosa la validez de los caracteres diagnósticos utilizados. Aún sabido esto, varios autores, quizás con una falta de visión
global o con una marcada falta de objetividad ante otras opiniones, siguen esta pauta y se aferran a la defensa a ultranza
de alguna de sus propias especies posteriormente sinonimizadas, con argumentos más subjetivos y “afectivos” que científicos,
y lo que es más grave, ignorando o utilizando más el demérito, el desprecio, la injuria o la descalificación de trabajos y
autores previos que la objetividad y humildad científicas. El tiempo, la Ciencia y su método traerán la opinión razonada sobre
estos procederes, y numerosos taxa deberán ser restudiados y redefinidos, teniendo en cuenta el valor real de los caracteres
utilizados y la potencial variabilidad entre las poblaciones dentro de sus áreas de distribución, y sin duda, el número y
posición de algunos taxa se verá modificado en el futuro, especialmente cuando se utilicen para su diagnosis los caracteres
genitales adecuados y permanentes y su variabilidad en aquellas especies de amplias distribuciones, y es seguro que nuevas
sinonimias serán de nuevo propuestas.
Por último anotar que, obviamente, a lo largo del pasado y el presente siglo se han ido publicando multitud de artículos y
revisiones relativas a faunas de diferentes países y/o continentes, algunas de las cuales hemos referido anteriormente, pero
también se han publicado centenares de publicaciones relacionadas con su Citología/Histología, Anatomía, Fisiología, Comportamiento,
Biogeografía, etc., y especialmente en relación con la Entomología Aplicada y/o con las faunas locales, que escapan a la intención
de esta contribución. Teniendo en cuenta la información general existente sobre esta familia, pasemos a centrarnos en la fauna
de la Península Ibérica y Baleares, motivo de esta contribución, y empecemos por su progresivo conocimiento en ella, motivo
que desarrollamos en el siguiente apartado.
SOBRE SU CONOCIMIENTO EN LA PENÍNSULA IBÉRICA Y BALEARES. PROBLEMÁTICA DE LAS CITAS ANTIGUAS
Como era de esperar en unos insectos tan pequeños y tan poco “vistosos”, las primeras citas en la Fauna Ibérica aparecerán
bastante más tarde que en el caso de otras familias con especies más llamativas o de mayor tamaño (Ascalaphidae, Nemopteridae,
Myrmeleontidae, incluso Chrysopidae, Hemerobiidae, etc.), y este grupo de insectos despertó mucha menos atención entre los
entomólogos y aficionados de las épocas iniciales y, consecuentemente, no tratamos un grupo de insectos frecuentemente citado
en la bibliografía de este periodo. Desde finales del siglo XIX y durante los dos primeros tercios del siglo XX, y al margen
de la extensa obra de L. Navás, autor del que ahora hablaremos, diferentes entomólogos han citado material ibérico o han hecho
referencia a alguno de los países de la Península Ibérica al citar la distribución general de algunas especies (McLachlan,
1868, 1880; Dziędzielewicz, 1905; Enderlein, 1906, 1930; Macho Bariego, 1909; Klapálek, 1909; Beché, 1913; Bohigas & Sanchez, 1917; Lacroix, 1924; Esben-Petersen, 1931; Killington, 1936; Eglin, 1940; Vidal y López, 1943; Janetschek, 1957; Tjeder, 1957; Zelený, 1961a, 1971; Berland, 1962; Kis, 1969, etc.), pero como ocurre en la bibliografía neuropterológica general y según hemos indicado, en mucha menor medida a lo acaecido
para otras familias de neurópteros más vistosas, de mayor tamaño o más “manejables”, y por citar un ejemplo Pictet (1865) no los recoge entre los neurópteros en su Synopsis des Névroptères d´Espagne, y que dejando para más adelante a L. Navás, ya enlazan con los autores más contemporáneos. Para los lectores que deseen
información y bibliografía sobre todos estos autores españoles (entre 1758-1990) se recomienda Martín Albaladejo (1994) y para los autores nacionales o no, que se interesaron en los neurópteros ibéricos se recomienda Monserrat & Triviño (2013).
Centrándonos más en lo que nos compete, y al margen de los citados autores del siglo XIX-primera mitad del XX que puntualmente
han aportado algún registro de especies de esta familia en nuestra fauna y que anteriormente hemos citado, no sería posible
concebir el estudio de los neurópteros en general, y de los coniopterígidos ibéricos (incluidos los de Andorra, España y Portugal)
en particular, sin las numerosas publicaciones del autor español Longinos Navás (1858-1938), prolífico, heterogéneo y controvertido
autor en numerosos campos, quien durante casi cuarenta años centró la mayoría de sus estudios en la taxonomía y sistemática
de este grupo de insectos, a quien debemos la mayoría de la información bibliográfica existente y la inmensa mayoría de las
citas del primer tercio del pasado siglo correspondientes a nuestra fauna, y fue él quien realizó la primeras revisiones de
los coniopterígidos incluidos en sus sinopsis de los neurópteros de España y de Cataluña (Navás, 1908, 1924a, 1925a), y que en el caso de la familia que nos ocupa, y por su extensión, no hemos pormenorizado en la relación de autores
y obras anteriormente citada, sino que la mencionamos en cada una de las especies estudiadas al referirnos a sus especies,
sinonimias y a la multitud de citas de coniopterígidos que este autor anotó para nuestra fauna y, consecuentemente, recogemos
sus artículos en la bibliografía que anotamos. Por su relevancia en nuestro estudio, es un autor que merece comentarse y en
él ahora nos detenemos.
Como sacerdote y católico Navás fue “lógicamente” un buen creacionista. Piénsese que sólo de estos tres órdenes de insectos
(Neuropterida) describió aproximadamente 375 géneros, 2.173 especies y 301 variedades (Monserrat, 1986), es decir 2.849 taxa, del total de 388 géneros y 2.684 especies por él descritos de diferentes órdenes, principalmente de
insectos y arácnidos (Monserrat, 1985c, 1986, 2011; Bastero Monserrat, 1989; Aspöck & Aspöck, 2014), y en lo que a nuestra fauna incumbe, de todos estos taxa, multitud de géneros, y nada menos que 152 especies y 149 variedades
de Neuropterida fueron descritas por él en la Fauna Ibero-Balear, que sumadas a las descritas de las Islas Canarias alcanzan
un total de 239 taxa: 4 de Megaloptera y 208 de Neuroptera (+ 15 de Canarias).
Como hemos citado anteriormente en el caso de otros entomólogos del pasado siglo, tampoco los coniopterígidos eran para Navás,
y nunca mejor dicho, “santo de su devoción” (“insectos muy pequeños”; “pequeñitos”; “alas cubiertas con escamitas”, etc.,
cita sobre ellos a lo largo de su obra), y no prestó demasiado interés en este grupo de insectos (Monserrat, 1984a), donde únicamente los cita en 38 obras y artículos de su extensa bibliografía neuropterológica ibérica (Monserrat & Triviño,
2013 recogen 171 obras y artículos suyos relacionados con nuestra fauna). Lamentablemente los criterios taxonómicos seguidos por
él han resultado ser de escaso valor y rigor científico, y sus identificaciones, citas y descripciones de nuevos taxa han
sido frecuentemente puestas en duda, ya que mayoritariamente en sus identificaciones no utilizaba el imprescindible estudio
de las estructuras genitales (“aparell sexual extern poc marcat” cita sobre ellos en su revisión de la Fauna de Cataluña,
Navás, 1924a) en la diagnosis e identificación de los ejemplares (sin tener en cuenta excelentes obras ya existentes, como las de Enderlein, 1906, 1908, que sin duda conocía y del que toma algunas imágenes, p. ej.: Navás, 1908), y basaba su criterio y sus estudios casi exclusivamente en caracteres como la venación o coloración alar, en gran parte
muy variables o sin verdadero interés (Withycombe, 1922a; Killington, 1929), y por ello sus trabajos consisten en un compendio de citas y taxa nuevos, que más que un avance, aportaron mucha confusión,
y por ello su obra ha sido permanentemente motivo de controversia, muy frecuentemente cuestionada, y afecta a la taxonomía
de esta familia y de los neurópteros en su conjunto. Como ejemplo, de los muchos que podríamos poner, comentemos que Mansell (2010) menciona 764 taxa del orden Neuroptera descritos por este autor de la Región Afrotropical (especies en su mayoría) de las
que el 50 % ya han sido consideradas no válidas, mayoritariamente sinonimizadas, y 388 taxa aún permanecen en posición incierta
o con su material tipo destruido o perdido en paradero desconocido. Como otros ejemplos de su quehacer, y en particular relacionado
con la familia que tratamos, citemos que Monserrat (1986) recoge 2 géneros, 17 especies y 1 variedad de coniopterígidos descritos por este autor a lo largo de su vida, Monserrat (1985c) revisa los tipos que quedaron de su casi destruida colección personal y solo los tipos de 6 especies y 1 variedad sobrevivieron,
Meinander (1986) refiere como sinónimas de especies previamente descritas todas sus especies descritas entre 1912-1918 de Norteamérica, y Monserrat (2011) propone como sinonimias de especies previamente descritas a dos de sus especies de nuestra fauna que hasta entonces se hallaban
en nomina dubia.
Como conclusión a su labor en la taxonomía de esta familia y en lo que nos compete, del género (Deasia) y las 5 especies de coniopterígidos descritas por Navás de la fauna ibero-balear (ver relación en el listado de taxa), casi todas han permanecido inicialmente en nomina dubia o han acabado siendo sinonimizadas (Meinander, 1972a; Monserrat, 2011; Monserrat & Triviño, 2013) y solo una (Coniopteryx parthenia) ha mantenido su validez por un tiempo, hasta que ha sido recientemente propuesta como sinonimia (Günther, 1993; Aspöck & Aspöck, 2001, 2014). Curiosamente dos de sus especies han acabado siendo revalidadas (por el mero Principio de Prioridad): una (Helicoconis interna), que ha sido recientemente revalidada (Monserrat, 2011), y ahora discutimos y proponemos la validez de otra (Coniopteryx parthenia).
Desafortunadamente casi son solo sus publicaciones las que nos quedan, ya que su colección privada sufrió diversas y penosas
vicisitudes a lo largo de los años (Monserrat, 1985c, 1986), y por tanto, los ejemplares con los que contamos actualmente depositados en instituciones para revisar su labor son proporcionalmente
escasos. Sobre este particular, y en lo que respecta al legado de Navás sobre esta familia y a nuestra faunística histórica,
nuevos comentarios fueron dados en base a nuevos elementos y datos sobre nuestra fauna (Ohm, 1965; Meinander, 1972a; Aspöck et al., 1980; Monserrat, 1984g, 1985d, 1995; Monserrat & Díaz-Aranda, 1990, etc.), pero en particular citemos a Monserrat (1984a) quien estudia los coniopterígidos que de su colección personal (hoy bajo la custodia del Museo de Ciencias Naturales de
Barcelona) pudieron salvarse (142 ejemplares), y es tal la cantidad de errores de identificación encontrados (prácticamente
todos los ejemplares estaban mal identificados, hecho absolutamente esperable, dado que no utilizaba la genitalia para las
identificaciones), que propone no considerar las citas faunísticas dadas por este autor, y que en el caso de nuestra fauna
son, en muchos casos, extremadamente frecuentes (ver listado de sus referencias en cada especie), afectando al número de especies
errónea-y a veces abundantemente citadas en la fauna ibérica por Navás (incluso generando numerosas referencias posteriores
de otros autores en la fauna que tratamos) y que no han sido corroboradas (ver apartado final de citas dudosas), y por ello
no quedan incluidas en la lista de especies ibéricas (p. ej. Coniopteryx pygmaea o Helicoconis lutea). Por tanto, el legado de Navás ha quedado reducido realmente a alguna especie descrita que ha mantenido su validez taxonómica
(ver listado de especies consideradas y sus sinonimias) y a ciertos datos sobre la potencial distribución geográfica de algunas
especies.
Para acabar con la faunística primeva, citar que igual que ocurre con otras muchas citas dadas por autores de la primera mitad
del pasado siglo que no identificaban sus especies en base a la genitalia, éstas han de tomarse con mucha cautela. El haber
recogido ahora estas reseñas en cada una de las especies de nuestra fauna ha supuesto un gran esfuerzo, pero en realidad es
una labor meramente enciclopedista y puramente testimonial, y en ningún caso (sin haber sido reestudiado el material), ha
de tomarse esta información como mínimamente fiable o fidedigna. Por todo lo anotado, con respecto a las citas y referencias
existentes en la bibliografía relacionada con la zona tratada, sólo han de considerarse las dadas más recientemente por autores
solventes que utilizan la genitalia para la identificación de las diferentes especies, y no deben considerarse fiables las
citas anteriores, ni pueden darse por válidas sin problema de considerarlas dudosas o erróneas. Aun así, y para recopilar
toda la información existente, anotamos las citas bibliográficas existentes hasta esa fecha de estas especies en la Península
Ibérica y Baleares, que han sido citada en ellas bajo diferentes sinonimias o combinaciones nomenclatoriales.
Al margen de estos datos históricos, y en lo que respecta a la Fauna Ibero-Balear, tras la obra de Navás se abre un largo
periodo de silencio en el progreso al conocimiento de este grupo de insectos en nuestra fauna, salvo las referencias a alguna
de sus citas y/o especies. Un poco más adelante, algunas referencias y novedades puntuales empiezan a aparecer (Ohm, 1965; Aspöck & Aspöck, 1965a, 1969), pero en cualquier caso el nivel de conocimiento entonces era extraordionariamente precario. Baste mencionar que en la revisión
de la familia a nivel mundial, Meinander (1972a) únicamente hace referencia a alguno de los países ibéricos en solo 17 especies, de las que en 5 ocasiones se refiere a dudas
sobre la identidad o valía de especies descritas o citadas por Navás, de 11 especies simplemente se refiere a especies citadas
por los anteriores autores mencionados (o datos de ellos, bien in litt., o bien com. per.), y en solo 2 especies (C. parthenia y S. pseudouncinata) cita material ibérico.
Pero sin duda, tratamos la familia de neurópteros que más recientemente ha experimentado un mayor incremento en cuanto al
número de especies descritas y/o citadas en la fauna ahora considerada. Al margen de las especies históricas, recientemente
la familia ha experimentado en la zona de estudio un enorme auge, con multitud de descripciones de nuevas especies o de novedades
para la fauna ibero-balear (Aleuropteryx juniperi, Coniopteryx tineiformis, C. haematica, C. drammonti, C. arcuata, C. esbenpeterseni, C. lentiae, C tjederi,
C. borealis, C. loipetsederi, Semidalis vicina, Helicoconis hirtinervis), para la Fauna Europea (Hemisemidalis pallida, C. atlasensis, C. kerzhneri, C. atlantica, Aleuropteryx minuta, A. maculata, Parasemidalis triton,
Semidalis pluriramosa), o para la Ciencia (Nimboa espanoli, S. pseudouncinata, Helicoconis hispanica, H. iberica, H. panticosa, N. adelae, Aleuropteryx iberica, A. remane,
Coniopteryx perisi, C. ketiae, C. ezequi), representando la zona del continente más rica en especies de esta familia (Aspöck et al., 1980, 2001; Aspöck, 1992). De ellas, alguna era previamente conocida de Europa, presumiéndose su presencia en el área de estudio, pero
otras sólo eran conocidas de zonas norteafricanas o de zonas asiáticas muy alejadas, o resultaron nuevas para la Ciencia,
y que representaron interesantes novedades (ver lista de taxa), destacando, por el interés para nuestra fauna, las contribuciones
de Meinander, 1963, 1965b, 1972a, 1976, 1981, 1990, 1998a; Ohm, 1965, 1968, 1973a; Monserrat, 1975, 1976a, 1976b, 1977b, 1977c, 1978b, 1979c, 1980, 1981c, 1982b, 1984g, 1985d, 1987b; Rausch & Aspöck, 1978a; Rausch et al., 1978b; Aspöck et al., 1980, 2001; Monserrat & Díaz-Aranda, 1990; Monserrat & Triviño, 2013, etc. (Algunas de estas referencias, en principio, no afectaban a la fauna ibero-balear, pero en ellas se describieron especies
que posteriormente han sido halladas en ella). Desde el punto de vista de la ecología y los sustratos vegetales preferidos
por nuestras especies, destaca F. Marín (Marín & Monserrat, 1987, 1991a; Marín, 1994) con los primeros estudios ibéricos sobre comunidades de coniopterígidos, bien a nivel de determinados ecosistemas o bien
de una circunscripción provincial, y Monserrat & Marín (1992, 2001) compendiando toda la información existente hasta entonces sobre su preferencia de sustrato vegetal de las distintas especies
de nuestra fauna.
Recientemente se ha venido aportando en nuestra fauna abundante información sobre esta familia (ver referencias en cada especie),
aunque aún existen multitud de lagunas y áreas completamente inexploradas sobre este grupo, especialmente en Baleares y Portugal,
de hecho Monserrat & Triviño (2013) anotan que sólo un 57,55 % de la superficie ibérica y balear ha sido muestreada con el específico fín de recolectar neurópteros
(y Coniopterygidae entre ellos), y que sólo en un 30,82 % de su superficie se han hallado estos insectos en aquella fecha,
porcentaje que se ha visto levemente incrementado con nuevas zonas muestreadas desde entonces. Estos autores anotaban el reciente
enorme incremento de registros en nuestra fauna durante el último tercio del pasado siglo y lo que llevamos del presente,
que en el caso de la familia que tratamos se contaba en esa fecha con 5.118 registros en la Península Ibérica y Baleares (20,1
% del total de registros de neurópteros entonces contabilizados), y que se correspondían con 10.415 ejemplares ya recolectados
y/o estudiados por nosotros y a material inédito aún no citado en la bibliografía (1.264 ejemplares), que se suman al nuevo
material no recolectado entonces y que, en conjunto, ahora incluimos en el nuevo material estudiado (1.800 ejemplares de 32
especies) y 229 ejemplares citados por diversos autores en la bibliografía (76 referencias bibliográficas, de ellas 44 corresponden
al autor de esta contribución), de los que hemos considerado el material revisado por nosotros o dados por autores fiables,
cifra que se ha visto incrementada desde entonces y que ahora seguimos aumentándolo con el nuevo material que citamos (incluyendo
material complementario de otras zonas o países) y que hacen un total de 12.215 ejemplares estudiados, que sumados a los datos
aportados por autores recientes, hacen un total de 12.444 ejemplares en los que basamos esta revisión.
La mayoría de las especies ibéricas, que aún permanecían sin conocerse su correcta y adecuada posición taxonómica, han sido
recientemente resueltas por Monserrat (2011), y de ellas, tan solo quedaba por resolver la identidad de una presunta nueva especie áptera/ braquíptera citada por Janetschek (1957) como Aleuropteryx sp., de Granada (cotas altas en Sierra Nevada, ladera norte del Veleta, 2400-2700 m), y que Monserrat (2011, 2013) y Monserrat & Triviño (2013) presumen que pudiera tratarse de Helicoconis, y como Aleuropteryx sp. / Helicoconis sp. aparecen en su listado de especies, y que tras varios años de muestreos en la zona, hemos conseguido dilucidar (Figs. 139-144, 158). También hemos propuesto revalidar a Coniopteryx tullgreni para diferenciarla de C. haematica y así resolver la identidad real de los ejemplares del Mediterráneo y sur europeo respecto al material del norte europeo
(Figs. 188-194, 297-300), y seguimos manteniendo nuestro criterio asumiendo los de Enderlein (1906) y Meinander (1972a, 1981, 1990) frente a los de Günther (1993), H. Aspöck et al. (2001) y Aspöck & Aspöck (2014) respecto a la identidad de Coniopteryx parthenia y C. pygmaea.
Como hemos citado anteriormente, recientemente Monserrat (2011) ha resuelto la posición taxonómica de la casi totalidad de las especies ibéricas pertenecientes a esta familia que hasta
entonces poseían una situación taxonómica dudosa o sin resolver, y Monserrat & Triviño (2013) anotaban la problemática existente sobre las citas antiguas y la posición taxonómica de algunas especies citadas en la Península
Ibérica, aportando la lista de las especies ibero-baleares de esta familia con sus sinonimias y diferentes combinaciones nomenclatoriales
bajo las cuales han sido citadas en la bibliografía, y en base a las citas fiables existentes y a material inédito que en
parte ahora citamos, aportan un cartografiado de la distribución de las especies ibéricas, poniendo en evidencia que existen
enormes áreas aún no prospectadas, y que es de esperar que nuevas sorpresas nos depare el futuro. Por último Monserrat et al. (2013) aportan numerosas de estas citas inéditas e incrementan el número de especies conocidas en las faunas ibérica y europea.
Con respecto a la fauna balear, Monserrat (2005a) pone en duda las citas de alguna especie de marcada distribución eurosiberiana, y lista y aporta nuevo material de las 10
especies presentes en este archipiélago (no existe, por el momento, ningún endemismo conocido), y en relación a las Islas
Canarias puede consultarse McLachlan (1882a), Brauer (1900), Enderlein (1906, 1929, 1930), Navás (1906, 1912, 1916d), Withycombe (1925a), Klingstedt (1936), Meinander (1962b, 1965b, 1972a, 1990), Ohm (1963, 1986), Aspöck & Aspöck (1965a), Kis (1969), Monserrat & Reviejo (1977, 1978), Monserrat (1978c, 1978d, 1979b, 1984a, 1995), Ohm & Hölzel (1984, 1999), Domingo-Quero et al. (2003), Ohm & Báez (2004), Sziráki (2004, 2011), Izquierdo Moya et al. (2010) y Báez & Oromí (2010).
Obviamente también existen en nuestra fauna citas indeterminadas: Coniopteryx sp., Aleuropteryx sp., Helicoconis sp., Conwentzia sp., Nimboa sp., etc. (Janetschek, 1957; Meinander, 1972a; Monserrat, 1977a, 1978a, 1978b, 2011; Ribeiro, 1997; Letardi, 2012; Monserrat & Triviño, 2013, etc.) y que, salvo se hayan asignado posteriormente a alguna especie en concreto, no pueden ser consideradas. Por último,
y dado su carácter aplicado en silvicultura y especialmente en la agricultura, los coniopterígidos han sido también citados
de forma general o inespecífica en multitud de artículos de índole agronómico, sea sobre olivos (Ruiz & Montiel, 2001), cítricos (Ripolles & Melia, 1980; García Marí et al., 1983; García Marí & González Zamora, 1999; Martins et al., 2002; Soler et al., 2002; Alvis et al., 2003), etc., y un sin fin de datos relacionados con la agricultura que mayoritariamente escapan a la intención de esta contribución.
Ante todo ello, y dada la multitud de artículos faunísticos y de otra índole que han aportado numerosos datos sobre la taxonomía,
la morfología, la corología, los estadios larvarios y la biología de las especies ibero-baleares, puede indicarse que en el
caso de la Fauna Ibérica, y para la parte española en particular, han acabado por conformar una de las familias mejor conocida
y más rica en especies dentro de la Fauna Europea, representando la fauna más abundante y diversa del Continente Europeo.
Al margen de la corología, sobre la biología de las especies ibéricas, y de entre toda esta información que se iba recabando,
se han venido describiendo los estadios larvarios de algunas especies que eran desconocidos, y que junto a los ya existentes
en la bibliografía se anotan en cada caso.
Como en muchas otras familias de neurópteros, muchas especies de coniopterígidos son excelentes bio-indicadores de la calidad
y grado de conservación del Medio Ambiente y sufren la alteración humana del medio, requiriendo su protección (New, 1991b, 1998; Gepp, 1999), y quizás conviene aprovechar esta oportunidad para mencionar un pequeño apunte sobre nuestra experiencia en la recolecta
de estos insectos en la Península Ibérica durante las últimas cuatro décadas, en las que, como hemos observado en otros grupos
de neurópteros, hemos percibido una franca regresión en sus poblaciones, con una progresiva disminución de sus efectivos dentro
de numerosas poblaciones respecto a lo que, no hace demasiado tiempo, antes obteníamos, incluso potenciando más recientemente
las unidades de esfuerzo, intensidad y número de investigadores para recolectarlos, y son un ejemplo más del silencioso e
imparable deterioro sobre la Naturaleza que las acciones del hombre están provocando (Tellería, 2013) sin que apenas sea perceptible, salvo que la edad de algunos entomólogos (como es el caso) lo aprecie, y lo que es más grave,
sin que, en muchos casos, las autoridades “competentes” hagan nada (o casi nada) por detener este lamentable deterioro. Esto
ya se ha manifestado en otras zonas, algunas muy distantes (y así lo comenta Zimmerman, 1957 y New, 1991b en la fauna de Hawaii), hasta el punto de que algunas especies forman parte de la lista de especies amenazadas o en peligro
de extinción (Gepp, 1981, 1983, 1999; Ohm, 1984; Saure & Gerstberger, 1991; New, 1991b, 1998; Devetak, 1992; Pröse, 1992; Tröger, 1993; Gepp et al., 1994; Duelli, 1994a; Röhricht & Tröger, 1998; Hölzel, 1999; Ivanov & Krivokhatsky, 1999, 2002, 2004; Jedlička, 2001; Pröse & Gruppe, 2003; Röhricht, 2004; Zelený, 2005, etc.). No existe, que sepamos, ninguna especie de coniopterígido protegido en nuestra fauna.
Como no podía ser de otra forma, también los coniopterígidos (Figs. 1-9) son actualmente objeto de atención en Internet (McEwen & Oswald, 1998; Röhricht, 2002; Letardi, 2002a, 2002b, 2010, etc.), formando parte o siendo protagonistas en multitud de webs informativas, didácticas o académicas, y especialmente
lo son en multitud de foros que recientemente han aparecido, y por la misma razón aducida para las citas bibliográficas de
estas especies, estos registros, en particular los que nos competen, podrían considerarse siempre que los datos aportados
en estas webs sean los correctos, y en este caso muy pocas veces hay visu posible en las identificaciones, por lo que han de tomarse con mucha cautela, de hecho, y sin ánimo de demeritar la envidiable
vocación de muchos aficionados y participantes en estas redes, sorprende que en muchos de estos foros nuestras especies aparezcan
“correctamente identificadas”, así “a ojo”, como si fueran cromos de una colección infantil, hecho que nos parece sorprendente
y datos que, obviamente, han de tomarse con cautela.
A pesar de toda la información publicada hasta la fecha, y en lo que respecta a la Fauna Ibero-Balear, la familia Coniopterygidae
no ha sido objeto de una investigación profunda basada en todo el estudio del material del que disponemos en la actualidad.
Por otra parte, toda la información existente sobre este orden de insectos en la Península Ibérica y Baleares está muy dispersa
o aún es desconocida, y consideramos que es de gran importancia subsanar dicha laguna efectuando una revisión completa. En
cualquier caso, compendiar toda esta información, actualizar, revisar y contribuir al mayor conocimiento de esta familia en
la Península Ibérica es pues uno de los objetivos del presente artículo.
En ella se realiza una revisión de la familia Coniopterygidae de la Península Ibérica e Islas Baleares, aportando el listado
taxonómico actualizado de las especies conocidas en la zona, la recopilación de toda la información bibliográfica existente
relacionada con sus especies, sobre sus sinonimias comúnmente aceptadas y sobre las diferentes combinaciones nomenclaroriales
bajo las cuales han sido citadas en la bibliografía (siempre nos quedará alguna en el “tintero”), aportando el listado actualizado
de las especies conocidas en la zona, así como una clave de identificación de sus géneros (larvas e imagos) y de sus especies
(imagos), y de cada una de ellas, anotamos, cuando sea preciso, datos sobre su taxonomía, morfología y/o variabilidad, así
como los datos conocidos sobre sus estadios juveniles, cuando existan, y toda la información existente sobre su biología,
distribución geográfica, fenológica y altitudinal, tanto en general para cada especie, como referente a los datos conocidos
en nuestra fauna.
Basamos esta revisión en 10.415 ejemplares ya previamente recolectados y estudiados por nosotros, sumados a los 229 ejemplares
citados en la bibliografía, de los que hemos considerado el material revisado por nosotros o dados por autores fiables, y
aportamos nuevo material inédito ahora estudiado (1.800 nuevos ejemplares de 32 especies), que contribuye a incrementar su
conocimiento en nuestra fauna, y que hacen un total de 12.215 ejemplares estudiados, que sumados a los datos aportados por
autores recientes, hacen un total de 12.444 ejemplares en los que basamos esta revisión.
Por último, anotamos en un apartado final con alguna especie que consideramos incorrecta- o imprecisamente citadas en nuestra
fauna.
Material y MétodosTOP
Para la realización de esta revisión recopilamos y partimos de toda la información general existente en la bibliografía referente
a los coniopterígidos que han resultado de nuestro interés (760 referencias bibliográficas consultadas y citadas), y en el
caso de las especies de la fauna ibérico-balear, se han tenido en cuenta todos los taxa citados en nuestra fauna a partir
de toda la información bibliográfica existente sobre ella (76 referencias bibliográficas, de ellas 44 corresponden al autor
de esta contribución).
Conocida la relación de especies presentes en la fauna tratada, se anota la relación taxonómica y sistemática de los taxa
que nos competen. Los taxa y sus referencias bibliográficas se exponen tanto bajo la denominación nomenclatural actualmente
aceptada, como bajo la denominación original, las sinonimias habitualmente reconocidas u otras combinaciones nomenclaturales
en las que han sido citadas en la bibliografía ibérica.
Posteriormente se aporta una clave de identificación para los imagos de las subfamilias, géneros y especies (tanto de machos
como, hasta donde es conocido y nos ha sido posible, de hembras), y una clave a nivel genérico de sus fases larvarias conocidas.
Para la realización de estas claves aportamos imágenes de las alas de todos los géneros (Figs. 85-92) y de muchas de sus especies, junto a caracteres de genitalia, morfología y/o pigmentación tegumentaria necesarios para la
identificación específica, junto a ciertos datos sobre su biología, preferencia de sustrato vegetal o distribución que pueden
ayudar puntualmente en este cometido. Para comodidad del lector y para facilitar su detección en las figuras aportadas, en
algunos casos señalamos con flechas y/o letras los elementos morfológicos que referimos y así señalamos e intentamos destacar,
y en la morfología alar (Figs. 23, 24) y genital anotadas, indicamos para cada uno de los géneros los elementos y terminología que son habitualmente utilizados.
De cada una de las especies se anota la referencia de su descripción original (haya sido realizada en base a material de nuestra
fauna o no) y los datos que sobre nuestra fauna existen de cada una de las especies tratadas (siempre se nos ha podido quedar
alguna en el “tintero”), recopilando todas las referencias bibliográficas existentes, e indicando las páginas donde se puede
recabar la información aportada sobre ellas. Las anotamos, tanto en la denominación actualmente aceptada, como bajo los diferentes
nombres, sinonimias y combinaciones nomenclaturiales bajo los que han sido citadas. Se aportan, cronológicamente ordenadas,
las referencias existentes con los autores de estas citas, su fecha de publicación y la/s página/s donde en ellas se aporta
información sobre su biología (bio), descripción original (d), distribución geográfica (dis), fenología (fen), morfología
del huevo (h), inclusión en claves (key), larva (l), aparezca en un listado (list), morfología del imago (mf), pupa (p), simple
referencia a alguno de los países de la Península Ibérica (rf), o datos sobre su taxonomía (tx). En las obras de carácter
general, solo se han anotado los datos referentes al material ibero-balear en ellas citado.
En principio, y particularmente en lo referido a la mayoría de las citas antiguas, esta recopilación anotada es meramente
informativa, y no implica la total asunción de sus contenidos, ni su vinculación taxonómica real. Por todo ello, para evitar
la utilización de datos poco fiables citados en la bibliografía y correr el riesgo de asumir como ciertos datos basados en
identificaciones posiblemente erróneas o incorrectas, mezclando información de unas y otras especies, y habida cuenta del
abundante material estudiado en las cuatro últimas décadas (10.415 ejemplares), para la realización de este estudio sobre
la biología, fenología y distribución altitudinal y fenológica de las especies ibérico-baleares, sólo se ha tenido en cuenta
este material revisado, estudiado y ya publicado por nosotros, y puntualmente los datos aportados por autores recientes de
reconocida solvencia (229 ejemplares), que se suman al nuevo material ahora citado y anotado para 32 especies (1.800 ejemplares)
y que, como hemos indicado, hacen un total de 12.444 ejemplares en los que basamos esta revisión.
Cuando lo hemos considerado necesario, citamos los datos más destacables sobre su taxonomía, morfología externa y/o variabilidad,
y para las especies de identidad más problemática o con citas cuestionables y/o dudosas en nuestra fauna se anota un apartado
final donde se aportan los datos que hemos considerado necesario comentar.
En lo que respecta al nuevo material ahora estudiado, se corresponde con material inédito por nosotros estudiado y recolectado
en diferentes puntos de la Península Ibérica y Baleares, mayoritariamente perteneciente a la colección del autor (Universidad
Complutense, Madrid, España), así como aquel otro material perteneciente a ciertas instituciones y colegas que amablemente
nos han permitido el estudio de sus colecciones o que nos han proporcionado información sobre el tema que nos ocupa y cuyas
siglas se anotan a continuación:
| AG: |
Colección Dr. Axel Gruppe (Freising, Alemania). |
| DV: |
Colección Daniel Ventura (Menorca, España). |
| IFAPA: |
Instituto Andaluz de Investigación y Formación Agraria, Laboratorio de Entomología Agraria, Cortijo de la Cruz (Málaga, España). |
| MNCN: |
Museo Nacional de Ciencias Naturales (Madrid, España). |
| MG: |
Museo Civico di Storia Naturale Giacomo Doria (Génova, Italia). |
Para la exposición del nuevo material estudiado, los datos se anotan alfabética- y cronológicamente ordenados, indicándose,
en función de los datos de captura disponibles y por este orden, el país, provincia-isla y localidad de captura, coordenadas
UTM o geográficas, altitud en msnm, fecha de captura, número de ♂♂, de ♀♀, o de estadios juveniles estudiados, planta sobre
la que los ejemplares han sido capturados (por economía de espacio, se han apocopando los nombres de los géneros cuando ya
ha sido utilizado), recolector (por igual motivo, únicamente se indica el recolector cuando no ha sido el autor de esta contribución)
e institución a la que pertenecen según las siglas anotadas anteriormente (por igual motivo, solo en el caso de que el material
pertenezca a alguna de ellas y no a la colección del autor). Para aportar nuevos datos sobre las especies del área estudiada,
en algunos casos también se anota, como material complementario de comparación, otro material no ibérico estudiado.
Para cada una de las especies se comentan los datos generales conocidos más destacables sobre ellas, sobre su morfología externa,
segmentos genitales, biología, fases juveniles (en su caso conocidas) y distribución fenológica, altitudinal, y geográfica
en la zona, hábitat y plantas soporte sobre las que han sido mayoritariamente recolectadas en base a los datos conocidos y
corroborados para la fauna tratada, así como, en algún caso, un apartado de discusión, donde se anotan los datos que hemos
considerado necesario comentar. Para la distribución altitudinal recabada para cada especie también se ha considerado 10 m
como cita/recolecta a nivel del mar, y para las fenologías de cada especie se anotan en números romanos para indicar los meses
en los que han sido recolectado los imagos, y con indicación de interrogación si algún dato es dudoso, no habiendo sido considerados
los datos de los ejemplares obtenidos ex ovo y/o cultivados ex larva artificialmente en condiciones no naturales.
Aunque en algunos casos existen datos en la morfología externa que ayudan a la identificación de algunas especies ibéricas
de esta familia, y así los utilizamos en las claves, el estudio de la genitalia es absolutamente necesario para la segura
y correcta identificación de los ejemplares. Para ello se procedió a la separación de los últimos cuatro segmentos del abdomen
mediante disección. Si el material estaba conservado en seco se ha hidratado previamente durante 24 horas por inmersión en
agua, y posteriormente se han aclarado estos últimos segmentos con ácido láctico caliente o hidróxido potásico al 10 % templado
durante el tiempo necesario para acceder a las estructuras genitales. Para las hembras se recomienda una leve tinción con
negro de clorazol (1 %), o bien con eosina, hematoxilina o mercuriocromo. Una vez realizado este proceso hemos introducido
la genitalia en glicerina dentro de un portaobjetos excavado para su observación mediante un estereo-microscopio ZEISS® STEMI-SV8
con cámara clara de dibujo incorporada, que también ha sido utilizado para la observación, identificación y descripción de
la mayoría de los ejemplares. Los dibujos se realizaron a mano alzada con cámara clara, pasándose posteriormente a papel vegetal
y tinta china, y tras escanearse se retocaron utilizando el programa de diseño Corel Paint Shop Pro Photo XI, y el software
Adobe Photoshop CS5 Extended Version 12.0 fue utilizado para el procesamiento de alguna de las imágenes.
Sobre la taxonomía, sistemática y morfología general se ha elegido la terminología habitualmente usada para esta familia (Meinander, 1972a; Aspöck et al., 1980), si bien la terminología de la venación alar en esta familia es objeto de diversas interpretaciones (Enderlein, 1906, 1908; Withycombe, 1922b; Killington, 1936; Kis et al., 1970; Grimaldi et al., 2013, etc.), habiendo adoptado la de los autores anteriormente citados (Figs. 23, 24). Para la distribución general de las especies paleárticas occidentales y/o europeas de la familia se recomienda la recopilada
por H. Aspöck et al. (2001), y sobre la preferencia de sustrato vegetal de las especies citadas puede consultarse Aspöck et al. (1980), y particularmente en nuestra fauna Monserrat & Marín (1992, 2001).
ResultadosTOP
LOS CONIOPTERÍGIDOS DE LA PENÍNSULA IBÉRICA Y BALEARES
Listado de subfamilias, tribus, géneros, subgéneros y especies considerados:
Para la taxonomía, en este listado sólo se consideran los taxa que afectan a la fauna considerada. En cada una de las especies
que tratamos se citarán las sinonimias habitualmente reconocidas y las diferentes combinaciones nomenclatoriales con las que,
directa- o indirectamente han sido citadas en la Fauna Ibérica y Balear.
El orden de los géneros dentro de cada categoría superior no implica mayor o menor posible relación entre ellos, y tampoco
el de las especies dentro de cada género, que para comodidad del lector, están mayoritariamente ordenadas alfabéticamente.
CONIOPTERYGIDAE Burmeister, 1839
Aleuropteryginae Enderlein, 1905
Género tipo: Aleuropteryx Löw, 1885
Tribu Aleuropterygini Enderlein, 1905
Género tipo: Aleuropteryx Löw, 1885
Género Aleuropteryx Löw, 1885
Especie tipo: Aleuropteryx loewii Klapálek, 1894 (ver Oswald & Meinander, 1988 e ICZN, Opinion 1595, 1990).
Aleuropteryx iberica Monserrat, 1977
Aleuropteryx juniperi Ohm, 1968
Aleuropteryx loewii Klapálek, 1894
Aleuropteryx maculata Meinander, 1963
Aleuropteryx minuta Meinander, 1965
Aleuropteryx remane Rausch, Aspöck & Ohm, 1978
Tribu Fontenelleini Carpenter & Lestage, 1928
Género tipo: Fontenellea Carpenter & Lestage, 1928 (mantenido siguiendo el artículo 40 del CINZ).
Género Helicoconis Enderlein, 1905
Especie tipo: Coniopteryx lutea Wallengren, 1871
Helicoconis hirtinervis Tjeder, 1960
Helicoconis hispanica Ohm, 1965
Helicoconis iberica Ohm, 1965
Helicoconis interna (Navás, 1911)
Helicoconis panticosa Ohm, 1965
Helicoconis bachi n.sp.
Coniopteryginae Burmeister, 1839
Género tipo: Coniopteryx Curtis, 1834
Tribu Coniopterygini Burmeister, 1839
Género tipo: Coniopteryx Curtis, 1834
Género Coniopteryx Curtis, 1834
Especie tipo: Coniopteryx tineiformis Curtis, 1834
Subgénero Coniopteryx Curtis, 1834
Especie tipo: Coniopteryx tineiformis Curtis, 1834
Coniopteryx (Coniopteryx) borealis Tjeder, 1930
Coniopteryx (Coniopteryx) ezequi Monserrat, 1984
Coniopteryx (Coniopteryx) parthenia (Navás & Marcet, 1910) sensu Meinander, 1972a
Coniopteryx (Coniopteryx) tineiformis Curtis, 1834
Subgénero Holoconiopteryx Meinander, 1972
Especie tipo: Coniopteryx turneri Kimmins, 1935
Coniopteryx (Holoconiopteryx) drammonti Rousset, 1964
Coniopteryx (Holoconiopteryx) haematica McLachlan, 1868
Subgénero Metaconiopteryx Kis, Nagler & Mândru, 1970
Especie tipo: Coniopteryx esbenpeterseni Tjeder, 1930
Coniopteryx (Metaconiopteryx) arcuata Kis, 1965
Coniopteryx (Metaconiopteryx) esbenpeterseni Tjeder, 1930
Coniopteryx (Metaconiopteryx) lentiae Aspöck & Aspöck, 1964
Coniopteryx (Metaconiopteryx) tjederi Kimmins, 1934
Subgénero Xeroconiopteryx Meinander, 1972
Especie tipo: Coniopteryx loipetsederi Aspöck, 1963
Coniopteryx (Xeroconiopteryx) atlantica Ohm, 1963
Coniopteryx (Xeroconiopteryx) atlasensis Meinander, 1963
Coniopteryx (Xeroconiopteryx) kerzhneri Meinander, 1971
Coniopteryx (Xeroconiopteryx) ketiae Monserrat, 1985
Coniopteryx (Xeroconiopteryx) loipetsederi Aspöck, 1963
Coniopteryx (Xeroconiopteryx) perisi Monserrat, 1976
Género Nimboa Navás, 1915
Especie tipo: Nimboa guttulata Navás, 1915
Nimboa adelae Monserrat, 1985
Nimboa espanoli Ohm, 1973
Género Parasemidalis Enderlein, 1905
Especie tipo: Parasemidalis annae Enderlein, 1905
Parasemidalis fuscipennis (Reuter, 1894)
Parasemidalis triton Meinander, 1976
Tribu Conwentziini Enderlein, 1905
Género tipo: Conwentzia Enderlein, 1905
Género Conwentzia Enderlein, 1905
Especie tipo: Conwentzia pineticola Enderlein, 1905
Conwentzia pineticola Enderlein, 1905
Conwentzia psociformis (Curtis, 1834)
Género Hemisemidalis Meinander, 1972
Especie tipo: Parasemisemidalis pallida Withycombe, 1924
Hemisemidalis pallida (Withycombe, 1924)
Género Semidalis Enderlein, 1905
Especie tipo: Coniopteryx aleyrodiformis Stephens, 1836
Semidalis aleyrodiformis (Stephens, 1836)
Semidalis pluriramosa (Karny, 1924)
Semidalis pseudouncinata Meinander, 1963
Semidalis vicina (Hagen, 1861)
Clave de subfamilias y géneros ibero-baleares (imagos)
| 1 |
Alas anteriores con dos venillas transversales entre SR+Ma y Mp (Fig. 23). Alas posteriores con origen de SR+Ma muy cerca de la base del ala (Fig. 23). Galea con tres sub-segmentos |
Aleuropteryginae (2) |
| - |
Alas anteriores con una venilla transversal entre SR+Ma y Mp (Fig. 24). Alas posteriores con origen de SR+Ma no cerca de la base del ala (Fig. 24). Galea con un segmento (Fig. 14) |
Coniopteryginae (3) |
| 2 |
Alas posteriores con venilla radial contactando con SR+Ma tras su bifurcación (Fig. 86) |
Tribu Fontenelleini, Helicoconis |
| - |
Alas posteriores con venilla radial contactando con SR+Ma antes de su bifurcación (Fig. 85) |
Tribu Aleuropterygini, Aleuropteryx |
| 3 |
Alas posteriores reducidas (Fig. 90) |
Conwentzia |
| - |
Alas posteriores no reducidas (Figs. 87-89, 91, 92) |
4 |
| 4 |
Alas posteriores con vena M no bifurcada (Fig. 87) |
Coniopteryx |
| - |
Alas posteriores con vena M bifurcada (Figs. 88, 89) |
5 |
| 5 |
Ambas alas con SR+Ma no bifurcada distalmente (Fig. 88) |
Nimboa |
| - |
Ambas alas con SR+Ma bifurcada distalmente (Figs. 89, 91, 92) |
6 |
| 6 |
Alas posteriores con venilla entre CUp y Mp+CUa situada tras su bifurcación (Fig. 92) |
Semidalis |
| - |
Alas posteriores con venilla entre CUp y Mp+CUa situada antes de su bifurcación (Figs. 89, 91) |
7 |
| 7 |
Alas posteriores con venilla entre CUp y Mp+CUa perpendicular a ésta (Fig. 89) |
Parasemidalis* |
| - |
Alas posteriores con venilla entre CUp y Mp+CUa oblícua a ésta (Fig. 91) |
Hemisemidalis* |
| |
| * |
Aunque en todos los casos el estudio de la genitalia es imprescindible para la identificación específica, en estos dos géneros
debe recurrirse a ella para alcanzar con mayor seguridad el nivel de género, dado lo subjetivo y a veces variable de este
carácter. |
Clave de especies (imagos)
A diferencia de otras familias de neurópteros donde numerosos caracteres de morfología externa y de pigmentación tegumentaria
o alar pueden utilizarse para la identificación de los individuos, independientemente de su sexo (Myrmeleontidae, Nemopteridae,
Crocidae, Ascalaphidae, Chrysopidae, Sisyridae, Hemerobiidae, etc.), mayoritariamente la taxonomía en esta familia a nivel
de identificación específica está basada en la genitalia, tanto externa como interna, y mayoritariamente se ha utilizado la
masculina, más ocasionalmente la femenina (Aleuropteryx) cuando posee elementos bien esclerificados y visibles (Figs. 96, 102, 103, 105-107, 113-117, 133-138). Aunque la morfología de la genitalia femenina externa y ciertos elementos internos han sido descritos en otras ocasiones,
especialmente en algunos géneros (p. ej.: Coniopteryx) para diferenciar las especies comunes en un mismo país (Enderlein, 1906; Stitz, 1931; Tjeder, 1931, 1957; Killington, 1936; Meinander, 1962a, 1963, 1965a, 1965b, 1969a, 1969b, 1971, 1972a, 1972b, 1974b, 1974c, 1974d, 1975b, 1975d, 1977, 1979b, 1980, 1983a, 1983b, 1990, 1998b; Aspöck & Aspöck, 1965a; Monserrat, 1977a, 1977b, 1978d, 1981a, 1982c, 1984g, 1985c, 1989b, 1995, 1996b, 2005b; Rausch & Aspöck, 1978a; Rausch et al., 1978b; Aspöck et al., 1980; Meinander & Penny, 1982, etc.), también entre las especies europeas (Figs. 291-292), recientemente Sziráki (1992a, 1992b, 1992c, 1996, 1998a, 2002a, 2004; Wang et al., 2007, etc.) han iniciado el estudio de las estructuras genitales femeninas internas de numerosas especies, muchas de ellas europeas
(Figs. 293-315), que parecen tener sumo interés en la identificación de las hembras en numerosos géneros que carecen de otros caracteres
para su identificación y que habitualmente venían siendo asignadas a las especies de los machos junto a los cuales se recolectaban.
Estos nuevos elementos han aportado nueva luz para la correcta identificación de las hembras de varios géneros, especialmente
los que poseen un mayor número de especies (Coniopteryx, Semidalis) y que, debido a su naturaleza membranosa y pocas veces pigmentada/esclerificada, exige tinción y un estudio mucho más detallado.
Para las especies de genitalia femenina interna conocida o ahora aportada, se complementará la clave con elementos para las
hembras.
En algunos casos se anotan ciertos datos complementarios sobre la distribución y/o la biología (en particular la/s planta/s
soporte habitual/es) de algunas especies que ayudan y orientan a su identificación.
Género Aleuropteryx
Fácilmente identificables tanto los machos (pedicelo con un agudo proceso distal: Fig. 94) como hembras (sin él).
| 1 |
Alas anteriores con manchas oscuras (Figs. 85, 104, 112) |
2 |
| - |
Alas anteriores sin manchas oscuras (Figs. 23, 93)* |
3 |
| 2 |
Alas anteriores con CUp sinuosa según Fig. 112. Genitalia masculina y femenina según Figs. 113-120. Especie mayoritariamente asociada a enebros y sabinas en el centro y sudeste peninsular |
Aleuropteryx maculata |
| - |
Alas anteriores con CUp no sinuosa según Fig. 104. Genitalia masculina y femenina según Figs. 105-111. Especie mayoritariamente asociada a tarays/tamarindos en el sudeste árido peninsular |
Aleuropteryx minuta |
| 3 |
Especie fuertemente asociada a encinas, alcornoques y coscojas. Alas según Fig. 93. Genitalia masculina y femenina según Figs. 95-103 |
Aleuropteryx iberica |
| - |
Especies mayoritariamente asociadas a otro tipo de vegetación. Genitalia masculina y femenina según Figs. 121-138 |
4 |
| 4 |
Especie mayoritariamente asociada a zonas secas y térmicas, carentes de vegetación arbórea, del centro y sudeste peninsular.
Genitalia masculina y femenina según Figs. 129-132, 137-138 |
Aleuropteryx remane |
| - |
Especies mayoritariamente asociadas a coníferas en todo el territorio peninsular. Genitalia masculina y femenina según Figs. 121-128, 133-136 |
5 |
| 5 |
Genitalia masculina y femenina según Figs. 121-124, 133-134 |
Aleuropteryx juniperi |
| - |
Genitalia masculina y femenina según Figs. 125-128, 135-136 |
Aleuropteryx loewii |
| |
| * |
Muy ocasionalmente, algunos ejemplares de Aleuropteryx juniperi pueden portar sombras oscuras en las celdillas marginales de las alas anteriores (Fig. 1), incluso alguna más tenue en las alas posteriores, por lo que el estudio de la genitalia en ambos sexos es necesaria. Aún
así, sólo en A. maculata y A. minuta (Figs. 85, 104, 112) las manchas están definidas y aisladas, mientras que lo mencionado en ejemplares de A. juniperi, las manchas son mucho más difusas y extendidas (Fig. 1). |
|
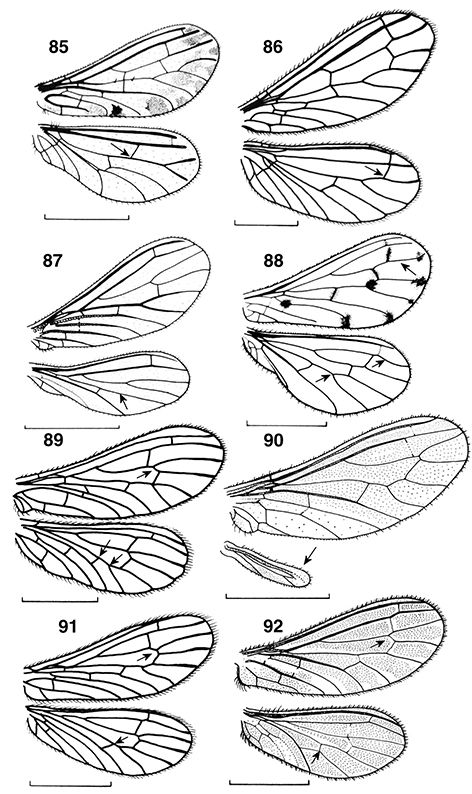 Figs. 85-92.— Venación alar de diferentes géneros de coniopterígidos: 85: Aleuropteryx, de Monserrat (1985d), 86: Helicoconis, 87: Coniopteryx, de Monserrat (1984g), 88: Nimboa, de Monserrat (1985d), 89: Parasemidalis, 90: Conwentzia, de Monserrat (1982c), 91: Hemisemidalis, 92: Semidalis, de Monserrat (1984f). Las flechas señalan algunos caracteres usados en las claves. Escala: 1 mm. Figs. 85-92.— Venación alar de diferentes géneros de coniopterígidos: 85: Aleuropteryx, de Monserrat (1985d), 86: Helicoconis, 87: Coniopteryx, de Monserrat (1984g), 88: Nimboa, de Monserrat (1985d), 89: Parasemidalis, 90: Conwentzia, de Monserrat (1982c), 91: Hemisemidalis, 92: Semidalis, de Monserrat (1984f). Las flechas señalan algunos caracteres usados en las claves. Escala: 1 mm.
Figs. 85-92.— Wings Venation in diferent dusty-wings genera: 85: Aleuropteryx, from Monserrat (1985d), 86: Helicoconis, 87: Coniopteryx, from Monserrat (1984g), 88: Nimboa, from Monserrat (1985d), 89: Parasemidalis, 90: Conwentzia, from Monserrat (1982c), 91: Hemisemidalis, 92: Semidalis, from Monserrat (1984f). Harrows indicates some characters used in keys. Scale: 1 mm.
|
|
Género Helicoconis
La subdivisión de este género en diferentes grupos de especies (Ohm, 1965), posteriormente elevados a categoría de subgéneros,
así como otros géneros posteriormente sinonimizados a él (Helicoconis s.str., Ohmopteryx, Fontenellea, Parahelicoconis, Capoconis) (Carpenter & Lestage, 1928; Kis et al., 1970; Meinander, 1972a; Sziráki, 2004; Tillier, 2013; Vidlička, 2014, etc.), no nos parece sostenible, concluyente, ni relevante en relación a los caracteres genitales utilizados en sus diagnosis
en otros géneros subdivididos en subgéneros (p. ej.: Coniopteryx), y que en ocasiones son de difícil asignación o están mezclados y/o se comparten según algunas especies. Por ello no han
sido tenidos en cuenta. Hasta el presente sólo los machos pueden identificarse (se anota aspecto genitalia femenina interna
en este género en Fig. 311).
| 1 |
Genitalia masculina con estilos portando un largo proceso interno basal (Fig. 145). Genitalia según Figs. 145, 146 |
Helicoconis hirtinervis |
| - |
Genitalia masculina con estilos sin un proceso interno basal (Figs. 140, 147, 152) |
2 |
| 2 |
Genitalia masculina con estilos marcadamente bifurcados en su extremo (Figs. 140, 141, 152, 157, 158) |
3 |
| - |
Genitalia masculina con estilos apenas o no bifurcados en su extremo (Figs. 147, 151) |
5 |
| 3 |
Genitalia masculina con parámeros portando una profunda incisión dorsal anterior a un fuerte dentículo (Fig. 153). Genitalia según Figs. 152-154 |
Helicoconis interna |
| - |
Genitalia masculina con parámeros sin portar una incisión dorsal ni un dentículo (Figs. 144, 155) |
4 |
| 4 |
Genitalia masculina con estilos largos y estilizados (Fig. 157). Parámeros rectos y estrechos (Fig. 155). Pene, en vista lateral, adelgazado hacia su extremo caudal de forma casi simétrica respecto a su eje, sin portar una globosa
formación membranosa ventral (Fig. 156) |
Helicoconis panticosa |
| - |
Genitalia masculina con estilos cortos y robustos (Figs. 140, 141, 158). Parámeros curvos y espatulados (Fig. 144). Pene, en vista lateral, adelgazado hacia su extremo caudal de forma asimétrica respecto a su eje, portando una globosa
formación membranosa ventral (Figs. 141, 143). Genitalia según Figs. 139-144, 158 |
Helicoconis bachi n.sp. |
| 5 |
Genitalia masculina con estilos no bifurcados en su extremo (Fig. 147). Pene portando dos procesos dorsales membranosos y extremo caudal agudo y no bifurcado (Fig. 149). Genitalia según Figs. 147-149 |
Helicoconis hispanica |
| - |
Genitalia masculina con estilos muy levemente bifurcados en su extremo (Fig. 151). Pene sin procesos dorsales y extremo caudal bifurcado (Fig. 150) |
Helicoconis iberica |
Género Coniopteryx
Fácilmente identificables los machos, tanto por su genitalia como por algunos caracteres de morfología externa, más tediosa
la identificación de las hembras, en algunas especies la genitalia no ha sido descrita aún. Anotamos algunos datos sobre sus
plantas soporte o su distribución geográfica que orienten y/o ayuden a la identificación de las hembras en la clave de las
especies cuya genitalia es conocida.
Machos:
| 1 |
Parámeros mucho más cortos que el pene. Entoproceso y el pene formando un bucle circular cerrado (Figs. 195, 197, 199, 201) |
Subgénero Metaconiopteryx |
| - |
Parámeros más largos que el pene. Entoproceso y el pene sin formar un bucle circular cerrado (Figs. 161, 181, 184, 208, 216, 223) |
2 |
| 2 |
Estilo unido a la parte basal del gonarco (Figs. 206, 213, 224, 231) |
Subgénero Xeroconiopteryx |
| - |
Estilo unido a la parte distal del gonarco (Figs. 161, 167, 170, 181, 189) |
3 |
| 3 |
Gonarco dividido en dos placas independientes y separadas en la linea caudal media (Figs. 162, 164) |
Subgénero Coniopteryx |
| - |
Gonarco no dividido en dos placas independientes y fusionadas en la linea caudal media (Figs. 182, 190, 191) |
Subgénero Holoconiopteryx |
Subgénero Metaconiopteryx
| 1 |
Bucle interno formado por el entoproceso y el pene de tamaño mucho menor, en vista lateral, que el hipandrio, no superando
el límite anterior del noveno segmento (Figs. 195, 199). Entoprocesos dilatados en su extremo (Figs. 195, 199). Parámeros situados anteriormente al resto de la genitalia interna (Figs. 195, 199) |
2 |
| - |
Bucle interno formado por el entoproceso y el pene de tamaño similar o mucho mayor, en vista lateral, que el hipandrio, superando
el límite anterior del noveno segmento (Figs. 197, 201). Entoprocesos no dilatados en su extremo (Figs. 197, 201). Parámeros situados ventralmente al resto de la genitalia interna (Figs. 197, 201) |
3 |
| 2 |
Extremo del entoproceso fuertemente dilatado distalmente en un proceso triangular (Fig. 199). Incisión media del hipandrio muy cerrada (Fig. 200). Genitalia según Figs. 199, 200 |
Coniopteryx (Metaconiopteryx) arcuata |
| - |
Extremo del entoproceso levemente dilatado distalmente en un proceso espatulado (Fig. 195). Incisión media del hipandrio abierta (Fig. 196). Genitalia según Figs. 195, 196 |
Coniopteryx (Metaconiopteryx) esbenpeterseni |
| 3 |
Bucle interno formado por el entoproceso y el pene superando ampliamente el límite anterior del octavo segmento (Fig. 201). Hipandrio fuertemente escotado en su margan lateral externo (Fig. 201). Genitalia según Figs. 201, 202 |
Coniopteryx (Metaconiopteryx) tjederi |
| - |
Bucle interno formado por el entoproceso y el pene apenas superando el límite anterior del noveno segmento (Fig. 197). Hipandrio levemente escotado en su margan lateral externo (Fig. 197). Genitalia según Figs. 197, 198 |
Coniopteryx (Metaconiopteryx) lentiae |
Subgénero Xeroconiopteryx
| 1 |
Pedicelo globoso, notablemente mayor que el escapo (Figs. 19, 22, 238, 239). Flagelómeros sin anillos de setas escuamiformes (Figs. 19, 238). Vértex con dos formaciones triangulares opuestas portando multitud de pequeñas setas (Figs. 15, 229, 240). Genitalia según Figs. 231-237 |
Coniopteryx (Xeroconiopteryx) perisi |
| - |
Pedicelo de similar tamaño que el escapo (Figs. 218, 204, 241). Flagelómeros con anillos de setas escuamiformes (Figs. 12, 204, 211, 218, 219). Vértex con pequeñas setas uniformemente distribuidas, sin generar dos formaciones triangulares opuestas de pequeñas setas
(Fig. 242) |
2 |
| 2 |
Setas escuamiformes de los flagelómeros dispuestas en dos anillos, uno distal y otro medial situado entre los anillos de setas
normales (Fig. 211). Genitalia según Figs. 213-216 |
Coniopteryx (Xeroconiopteryx) kerzhneri |
| - |
Setas escuamiformes de los flagelómeros dispuestas en dos anillos, ambos en posición distal (Figs. 12, 204, 219) |
3 |
| 3 |
Ectoprocto formado por dos amplias láminas triangulares o rectangulares (Figs. 213, 214) |
4 |
| - |
Ectoprocto formado por dos pequeñas láminas triangulares (Figs. 205, 206, 224, 225) |
5 |
| 4 |
Ectoprocto triangular (Figs. 220, 221). Parámeros no bifurcados en su extremo caudal (Fig. 223). Margen caudal del hipandrio con dos expansiones espatuladas (Figs. 221, 222). Genitalia según Figs. 220-223 |
Coniopteryx (Xeroconiopteryx) ketiae |
| - |
Ectoprocto rectangular (Fig. 231). Parámeros bifurcados en su extremo caudal (Fig. 237). Margen caudal del hipandrio sin dos expansiones espatuladas (Fig. 232). Genitalia como en Figs. 231-237 |
Coniopteryx (Xeroconiopteryx) atlasensis* |
| 5 |
Margen caudal del hipandrio con una formación triangular media, no hendida sobre su línea media (Figs. 224, 225, 228). Genitalia según Figs. 224-228 |
Coniopteryx (Xeroconiopteryx) loipetsederi |
| - |
Margen caudal del hipandrio con una formación trapecial media, marcadamente hendida sobre su línea media (Figs. 206, 207). Genitalia según Figs. 205-210 |
Coniopteryx (Xeroconiopteryx) atlantica |
| |
| * |
No existen diferencias significativas en la genitalia masculina respecto a C. (X) perisi, por lo que se han utilizado sus imágenes. |
Subgénero Coniopteryx
| 1 |
Parámeros progresivamente adelgazados hacia su ápice caudal, sin presentar procesos dorsales o ventrales en su zona distal (Figs. 170, 171). Procesos caudales que marginan la incisión media del hipandrio levemente bifurcados o de aspecto aserrado (Fig. 172). Genitalia según Figs. 170-172 |
Coniopteryx (Coniopteryx) pygmaea* |
| - |
Parámeros progresivamente adelgazados hacia su ápice caudal, pero presentando procesos dorsales o ventrales en su zona distal
(Figs. 163, 168, 174, 177). Procesos caudales que marginan la incisión media del hipandrio no bifurcados o de aspecto aserrado (Figs. 165, 168, 175, 178) |
2 |
| 2 |
Gonarco y especialmente sus ramas caudales (estoprocesos) robustas y anchas (Figs. 173). Parámeros con dos procesos dorsales (Fig. 174). Genitalia según Figs. 173-175 |
Coniopteryx (Coniopteryx) tineiformis |
| - |
Gonarco y especialmente sus ramas caudales (estoprocesos) delgadas y tenues (Figs. 161, 167, 176). Parámeros con un único proceso dorsal (Figs. 163, 168, 177) |
3 |
| 3 |
Parámeros con proceso dorsal robusto y fuertemente esclerificado, y proceso ventral levemente denticulado (Fig. 163). Genitalia según Figs. 161-166 |
Coniopteryx (Coniopteryx) ezequi |
| - |
Parámeros con proceso dorsal delgado o membranoso, y proceso ventral carente de tenues denticulaciones (Figs. 168, 177) |
4 |
| 4 |
Incisión media del hipandrio en forma de V abierta (Fig. 169). Porción distal de los parámeros larga y fuertemente recurvada (Fig. 168). Genitalia según Figs. 167-169 |
Coniopteryx (Coniopteryx) parthenia |
| - |
Incisión media del hipandrio en forma de U cerrada (Fig. 178). Porción distal de los parámeros corta y recta (Fig. 177). Genitalia según Figs. 176-178 |
Coniopteryx (Coniopteryx) borealis |
| |
| * |
No presente en nuestra fauna, pero incluida en esta clave para ilustrar elementos citados en el texto. |
Subgénero Holoconiopteryx
| 1 |
Estilo con su rama caudal dilatada en su extremo, portando un pequeño dentículo dorsal (Figs. 181, 183). Parámeros con un tenue proceso dorsal en su extremo caudal (Fig. 185). Genitalia según Figs. 181-187 |
Coniopteryx (Holoconiopteryx) drammonti |
| - |
Estilo con su rama caudal progresivamente adelgazada, no dilatada en su extremo, ni portando un dentículo dorsal (Fig. 189). Parámeros con un amplio proceso dorsal en su extremo caudal (Fig. 193). Genitalia según Figs. 189-193 |
Coniopteryx (Holoconiopteryx) haematica |
Hembras:
| 1 |
Gonapófisis laterales fusionadas en una única placa (Figs. 291B, C) |
Subgénero Xeroconiopteryx 2 |
| - |
Gonapófisis laterales independientes, no fusionadas en una única placa (Fig. 292B) |
3 |
| 2 |
Noveno esternito fuertemente esclerificado (Figs. 291A-E) |
Coniopteryx (Xeroconiopteryx) kerzhneri |
| - |
Noveno esternito no especialmente esclerificado |
Resto de especies |
| 3 |
Bolsa copuladora fuertemente esclerificada y pigmentada (Figs. 295-298) |
Subgénero Holoconiopteryx 4 |
| - |
Bolsa copuladora no esclerificada ni pigmentada (Figs. 293, 301-304) |
5 |
| 4 |
Bolsa copuladora triangular en vista ventral (Fig. 296) |
Coniopteryx (Holoconiopteryx) drammonti |
| - |
Bolsa copuladora espatulada en vista ventral (Fig. 298) |
Coniopteryx (Holoconiopteryx) haematica |
| 5 |
Espermateca alargada/cilíndrica, tubular o arriñonada (Figs. 312-315) |
Subgénero Metaconiopteryx 6 |
| - |
Espermateca con aspecto de globo/saciforme (Figs. 293, 304) |
Subgénero Coniopteryx 9 |
| 6 |
Porción distal de la espermateca fuertemente ensanchada (Fig. 312) |
Coniopteryx (Metaconiopteryx) arcuata |
| - |
Porción distal de la espermateca muy escasamente ensanchada (Figs. 313-315) |
7 |
| 7 |
Espermateca gruesa y corta, levemente arqueada (Fig. 315) |
Coniopteryx (Metaconiopteryx) esbenpeterseni |
| - |
Espermateca delgada y larga, irregularmente arqueada (Figs. 313, 314) |
8 |
| 8 |
Espermateca con un par de bucles angulosos, de longitud menor a 10 veces su máxima anchura (Fig. 314) |
Coniopteryx (Metaconiopteryx) lentiae |
| - |
Espermateca con numerosos bucles redondeados, de longitud mayor a 30-40 veces su máxima anchura (Fig. 313) |
Coniopteryx (Metaconiopteryx) tjederi |
| 9 |
Especie fuertemente asociada a enebros y sabinas |
Coniopteryx (Coniopteryx) ezequi |
| - |
Especies mayoritariamente asociadas a otras coníferas (Pinus, Laryx o Abies) o a planifolios |
10 |
| 10 |
Especie mayoritariamente asociada a Pinus, Larix o Abies. Bolsa copuladora en forma de embudo (Figs. 293, 294) |
Coniopteryx (Coniopteryx) parthenia |
| - |
Especies asociadas a planifolios. Bolsa copuladora en forma de saco (Figs. 301-304) |
11 |
| 11 |
Espermateca mucho mayor que la bolsa copuladora, tan alta como ancha (Fig. 304) |
Coniopteryx (Coniopteryx) pygmaea |
| - |
Espermateca menor o de similar tamaño que la bolsa copuladora, más larga que ancha (Figs. 301, 302) |
12 |
| 12 |
Ducto que conecta la espermateca y la bolsa copuladora corto y cónico (Fig. 302) |
Coniopteryx (Coniopteryx) borealis |
| - |
Ducto que conecta la espermateca y la bolsa copuladora largo y progresivamente cilíndrico (Fig. 301) |
Coniopteryx (Coniopteryx) tineiformis |
Género Nimboa
Tanto machos como hembras pueden identificarse sin problema merced a la pigmentación de las alas. Obviamente la genitalia
masculina es definitiva.
| 1 |
Alas anteriores manchadas (Fig. 243). Genitalia masculina según Figs. 244-249 |
Nimboa adelae |
| - |
Alas anteriores no manchadas (Fig. 250). Genitalia masculina y femenina según Figs. 251-255 |
Nimboa espanoli |
Género Parasemidalis
Hasta el presente sólo los machos son adecuadamente identificables.
| 1 |
Gonarco largo, recto y cilíndrico (Figs. 260, 261). Hipandrio con dos procesos cónicos a ambos lados de la línea media (Fig. 261). Genitalia masculina según Figs. 260, 261 |
Parasemidalis triton |
| - |
Gonarco más corto, sinuoso y cónico (Figs. 256, 257). Hipandrio sin dos procesos cónicos a ambos lados de la línea media (Figs. 256, 257). Genitalia masculina y femenina según Figs. 256-259 |
Parasemidalis fuscipennis |
Género Conwentzia
Aunque se han citado algunas leves diferencias entre ambas especies ibéricas, bien en la posición de ciertas venillas transversales
en las alas anteriores y/o en el color de las antenas y las patas (Enderlein, 1905b), estos caracteres son muy variables y han sido desestimados (Meinander, 1972a), sin embargo, el número de flagelómeros de las antenas parece mantenerse como carácter diferencial entre ambas especies
(Zelený, 1961b) y puede ayudar a la identificación especialmente en el caso de las hembras (♂♂/♀♀ Conwentzia pineticola: 32-36/ 30-35, Conwentzia psociformis: 38-43/ 36-43), aunque sólo los machos son identificables con mayor seguridad. Los datos de plantas soportes son también
muy útiles para la identificación de ambas especies.
| 1 |
Especie asociada con coníferas. En los machos el proceso distal del ectoprocto está marcadamente bifurcado en su extremo,
siendo sus ramas, especialmente la más dorsal, de similar longitud que la región basal de las que emergen (Fig. 263). En las hembras la bolsa copuladora como en Fig. 266 |
Conwentzia pineticola |
| - |
Especie asociada con planifolios. En los machos el proceso distal del ectoprocto no está bifurcado en su extremo, apenas siendo
dos tenues lobulaciones, y en cualquier caso son mucho más cortas que la región basal de las que emergen (Fig. 262). En las hembras la bolsa copuladora según Figs. 264, 265 |
Conwentzia psociformis |
Género Hemisemidalis
Especie única Hemisemidalis pallida (Figs. 267-274).
Tratamos una especie de genitalia masculina bastante variable, especialmente en el gonarco y en la extensión y límite del
margen caudal del hipandrio. En las Figuras 268-270 anotamos la variabilidad encontrada en diversos ejemplares ibéricos estudiados,
siendo el más habitual el representado en la Fig. 269.
Género Semidalis
Hasta donde conocemos, sólo los machos son identificables con seguridad. Algunos datos de hábitat pueden colaborar en la identificación,
especialmente de las hembras, no obstante se anota la genitalia interna conocida en las hembras de algunas especies.
Machos:
| 1 |
Margen caudal del hipandrio con dos pequeños procesos cónicos portadores, cada uno de ellos, de una larga seta (Figs. 275, 276). Especie limitada al sudeste árido peninsular. Genitalia masculina según Figs. 275-278 |
Semidalis pluriramosa |
| - |
Margen caudal del hipandrio de aspecto diferente pero sin presentar dos pequeños procesos cónicos portadores, cada uno de
ellos, de una larga seta (Figs. 279, 280, 283, 286). Especies del todo el territorio peninsular y/o Baleares |
2 |
| 2 |
Uncini bien desarrollados y están fusionados ventralmente formando una única pieza (Figs. 281, 282). Genitalia masculina según Figs. 279-282 |
Semidalis vicina |
| - |
Uncini apenas desarrollados, o si lo están, nunca están fusionados ventralmente, sino formando dos elementos independientes
(Figs. 283-288) |
3 |
| 3 |
Uncini apenas desarrollados (Figs. 283-285), parámeros con dos marcados dentículos dorsales, aplanados, muy agudos en su extremo y fuertemente recurvados hacia adelante
(Figs. 284, 285). Especie eurioica. Genitalia masculina según Figs. 283-285 |
Semidalis aleyrodiformis |
| - |
Uncini bien desarrollados, formando dos elementos unguiformes independientes a ambos lados de los parámeros (Figs. 286-288), éstos con dos tenues dentículos cónicos, pequeños y escasamente recurvados hacia adelante (Fig. 287). Especie mayoritariamente citada sobre coníferas. Genitalia masculina según Figs. 286-288 |
Semidalis pseudouncinata |
Hembras:
| 1 |
Porción ventral bajo las gonapófisis laterales prominente y triangular (Figs. 289, 290). Especie limitada al sudeste árido peninsular |
Semidalis pluriramosa |
| - |
Porción ventral bajo las gonapófisis laterales no destacada. Especies del todo el territorio peninsular y/o Baleares |
2 |
| 2 |
Ducto entre la espermateca y la bolsa copuladora muy corto (Fig. 309). Especie mayoritariamente citada sobre coníferas |
Semidalis pseudouncinata |
| - |
Ducto entre la espermateca y la bolsa copuladora muy largo (Fig. 310). Especie eurioica |
Semidalis aleyrodiformis |
Clave de larvas ibero-baleares (géneros conocidos)
La morfología antenal y la quetotaxia cefálica son de gran importancia, por lo que se recomienda, en la mayoría de los casos,
aclarar previamente los ejemplares con ácido láctico. Los caracteres que se anotan corresponden a fases larvarias maduras.
Para la información sobre las especies cuyas fases juveniles se conocían puede utilizarse Gepp (1984) quien aporta información sobre las especies cuyas larvas habían sido descritas y Monserrat (2001) que también aporta una clave de las larvas de todos los géneros entonces conocidas a nivel mundial, y lógicamente las aportaciones
posteriores, especialmente de Brucheiserinae (Sziráki & Flint, 2007; Sziráki, 2007a). Nosotros indicaremos en cada una de nuestras especies, la información existente sobre sus fases preimaginales conocidas.
| 1 |
Antenas truncadas en el ápice (Figs. 32, 38, 42). Labro cubriendo únicamente la parte basal de las mandíbulas (Figs. 32, 38). Tegumento con frecuentes setas escuamiformes (Fig. 80) |
Aleuropteryginae (2) |
| - |
Antenas adelgazadas en el ápice (Figs. 27, 39, 41, 43). Labro cubriendo completamente las mandíbulas (Figs. 28, 39, 40, 41). Tegumento con setas normales, no escuamiformes (Figs. 63, 81-83) |
Coniopteryginae (3) |
| 2 |
Setas clipeo-labrales y dorsales cortas y curvas (Figs. 36, 61). Escapo menor a un tercio de la longitud del flagelo (Fig. 62) |
Helicoconis |
| - |
Setas clipeo-labrales y dorsales largas y escasamente curvadas (Figs. 32, 38). Escapo mayor que la mitad de la longitud del flagelo (Figs. 32, 33, 38) |
Aleuropteryx |
| 3 |
Patas mucho más largas que la anchura del tórax. Flagelo más de seis veces la longitud del escapo (Fig. 41). Más de 10 setas irregularmente dispuestas sobre la zona central del clípeo-labro (Fig. 41) |
Conwentzia |
| - |
Patas tan largas o más cortas que la anchura del tórax. Flagelo menor de seis veces la longitud del escapo (Figs. 27, 39, 43). No más de 5 setas regularmente dispuestas sobre la zona central del clípeo-labro (Figs. 27, 37, 39) |
4 |
| 4 |
Longitud del flagelo casi dos veces la del escapo (Figs. 43, 53, 60). Setas clipeo-labrales dispuestas en una hilera media (Fig. 37, 56, 57) |
Hemisemidalis |
| - |
Longitud del flagelo más de dos veces la del escapo (Figs. 27, 39, 65, 66, 71, 72). Setas clipeo-labrales mayoritariamente dispuestas en dos hileras a ambos lados de la linea media (Figs. 39, 40, 73) |
5 |
| 5 |
Dos pares de setas clipeo-labrales situadas tras una media anterior (Figs. 39, 73) |
Semidalis |
| - |
Un par de setas clipeo-labrales situadas tras una media anterior (Figs. 27, 40) |
Coniopteryx |
LAS ESPECIES DE CONIOPTERYGIDAE DE LA PENÍNSULA IBÉRICA Y BALEARES
(El orden de las especies dentro de cada género es meramente alfabético o ha sido adecuado correlativamente a la secuencia
numérica elegida para las figuras usadas, y por ello no implica una mayor o menor posible relación entre ellas).
CONIOPTERYGIDAE
Aleuropteryginae
Aleuropteryx iberica Monserrat, 1977 (Figs. 23, 84, 93-103)
Monserrat, 1977b: 366 (d), 1978a: 67 (bio, dis); Aspöck et al., 1980: 9 (dis), 141 (bio, mf, tx, rf); Monserrat, 1980: 191 (bio, dis), 1982a: 81(bio, dis), 1984a: 146 (dis, tx), 1984b: 105 (bio, dis), 1984c: 160 (bio, dis), 1984d: 42 (bio, dis), 1984e: 174 (bio, dis), 1984g: 28 (bio, dis), 1985b: 88 (bio, dis), 1985d: 130 (bio, dis), 1985e: 75 (bio, dis); Díaz-Aranda et al., 1986a: 1125 (bio, dis), 1986b: 1142 (bio, dis); Monserrat, 1987a: 142 (bio, dis); Marín & Monserrat, 1987: 350-359 (bio, dis); Monserrat & Díaz-Aranda, 1987: 182 (bio, dis), 1988a: 92 (bio, dis); Díaz-Aranda & Monserrat, 1988a: 120 (bio, dis), 1988b: 226 (bio, dis), 1988c: 223 (bio, dis); Monserrat & Díaz-Aranda, 1989: 53 (bio, dis), 1990: 41 (bio, dis); Meinander, 1990: 3 (rf), 6 (rf); Monserrat et al., 1991: 108 (rf); Marín & Monserrat, 1991b: 182 (bio, dis); Monserrat & Marín, 1992: 282 (bio); Poggi, 1993: 420 (bio, dis); Marín, 1994: 260-297 (bio, dis, fen); Monserrat et al., 1994: 56 (bio, dis); Marín & Monserrat, 1995a: 43 (bio, dis), 1995b: 118 (bio, dis); Monserrat, 1995: 112 (bio, dis), 1996a: 14 (list); Meinander, 1998a: 24 (rf); H. Aspöck et al., 2001: 167 (rf); Monserrat, 2002: 5 (bio, dis); Sziráki, 2004: 4 (rf); Monserrat et al., 2013: 53 (bio, dis); Monserrat, 2013: 299 (rf); Monserrat & Triviño, 2013: 20 (list), 115 (dis).
Aleuropteryx boabdil Rausch & Aspöck, 1978
Rausch & Aspöck, 1978a: 9 (d, dis); Aspöck et al., 1980: 141 (tx); Meinander, 1990: 3 (rf); Monserrat & Triviño, 2013: 20 (list).
Aleuropteryx sp.
Monserrat, 1977a: 154, 158 (dis, nt).
Especie conocida de España, Portugal, sur de Francia y Marruecos. Está mayoritariamente citada sobre Quercus esclerófilos (encinas y alcornoques), entre V-VIII, en altitudes de hasta 2070 m. En nuestra fauna ha sido citada en toda
su región de influencia mediterránea, mayoritariamente sobre Quercus esclerófilos (encinas, alcornoques y coscojas), entre II, V-IX, en altitudes que oscilan entre 10-1900 m.
Los estadios preimaginales son desconocidos, solo el huevo ha sido descrito por Monserrat (1977b) (Fig. 84).
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Álava, Las Campas de Gabo, 605 m, 30TWN1541, 17.VI.2005, 1♂ sobre Quercus ilex, S. Pagola. Almería, Turrillas, 13.VI.2012, 1♂, 1♀ sobre Q. rotundifolia, V.Triviño. Badajoz, Mirandilla, 3.VII.2005, 1♀ sobre Q. rotundifolia, Valverde de Burguillos, 23.V.2012, 1♂ sobre Q. rotundifolia. Cádiz, Pinsapar de Grazalema, 3.V.2005, 2♂♂ sobre Q. rotundifolia. Cuenca, Beteta, 40°34’37.40’’N 2°02’33.10’’O, 1310 m, 27.VI.2009, 1♂ a la luz, Puerto Cabrejas, 40°03’16.98’’N 2°20’45.76’’O, 1150
m, 27.VI.2009, 1♀ sobre Q. rotundifolia. Badajoz, Fregenal de la Sierra, 19.VI.2012, 1♂ sobre Q. rotundifolia. Huelva, La Corte, 19.VI.2012, 1♂ sobre Q. rotundifolia. Lérida, Biosca, 41°50’11,51’’N 1°21’55,95’’E, 500 m, 19.VII.2009, 1♂, 2♀♀ sobre Q. rotundifolia, Talteüll, 41°49’09,24’’N 1°19’30,62’’E, 440 m, 19.VII.2009, 1♂, 3♀♀ sobre Q. rotundifolia. Madrid, Fuentidueña de Tajo, 28.VI.2009, 1♂ sobre Q. rotundifolia. Málaga, Los Alcornocales, 21.VI.2012, 1♂ sobre Q. suber. Toledo, Cuerva, 5.VII.2014, 1♂ sobre Q. rotundifolia.
Aleuropteryx juniperi Ohm, 1968 (Figs. 121-124, 133-134)
Ohm, 1968: 15 (d); Ohm & Remane, 1968: 218 (dis); Aspöck & Aspöck, 1969: 28 (rf); Ward, 1970: 75 (rf); Meinander, 1972a: 42 (rf); Henry, 1976: 195 (rf); Monserrat, 1977a: 22 (list), 154 (key, mf), 158 (bio, dis), 1977b: 372 (list, mf), 1978a: 67 (bio, dis), 1978b: 182 (bio, dis); Aspöck et al., 1980: 141 (bio, mf, tx, rf); Monserrat, 1982a:81 (bio, dis, mf), 1984a: 146 (dis, tx), 1984b: 105 (bio, dis), 1984c: 160 (bio, dis), 1984d: 42 (bio, dis), 1984e: 174 (bio, dis), 1984g: 28 (bio, dis), 1985b: 88 (bio, dis); Díaz-Aranda et al., 1986b: 1142 (bio, dis); Popov, 1986: 168 (rf); Díaz-Aranda & Monserrat, 1988a: 120 (bio, dis), 1988b: 226 (bio, dis), 1988c: 223 (bio, dis); Monserrat & Díaz-Aranda, 1988a: 92 (bio, dis), 1989: 53 (bio, dis), 1990: 40 (bio, dis); Meinander, 1990: 7 (dis); Marín & Monserrat, 1990: 227 (bio, dis), 1991b: 182 (bio, dis); Monserrat & Marín, 1992: 282 (bio); Monserrat et al., 1994: 56 (bio, dis); Marín & Monserrat, 1995a: 43 (bio, dis), 1995b: 118 (bio, dis); Monserrat, 1995: 112 (bio, dis), 1996a: 14 (list); Blasco, 2000: 98 (bio, dis); H. Aspöck et al., 2001: 167 (rf); Monserrat, 2002: 5 (bio, dis), 2005a: 73 (bio, dis); Zimmermann, 2005b: 37 (dis); Monserrat, 2013: 299 (rf); Monserrat et al., 2013: 53 (bio, dis); Letardi et al., 2013: 36 (list); Monserrat & Triviño, 2013: 20 (list), 115 (dis).
Especie Holomediterránea, también citada en Gran Bretaña y en Pennsylvania, Maryland, New Jersey, Michigan, New York, y Virginia
en USA (probablemente introducida por acción humana). Generalmente recolectada sobre Cupresaceae (Juniperus y Cupressus), pero también sobre otras coníferas, especialmente Pinus, y asimismo sobre muchas otras plantas (Quercus, Castanea, Ulmus, Ceratonia, Pistacea, Citrus, Olea, etc.), mostrando una alta versatilidad en su sustrato vegetal y ecológico. Está citada entre IV-VIII, en altitudes de 10
hasta 1500 m. En nuestra fauna ha sido ampliamente citada en toda su región de influencia mediterránea, incluyendo Baleares
(Formentera), citada sobre Juniperus, Cupressus, Pinus, Pistacea, Citrus, Castanea, Quercus esclerófilos o marcescentes, Ceratonia, Olea, etc., entre IV-IX, en altitudes que oscilan entre 10-2580 m.
Como hecho inusual en esta especie debemos comentar que algunos de los ejemplares ahora citados (especialmente ♀♀) recolectadas
en Granada (España), en Dragunara (Cerdeña, Italia) y alguno de los que ahora representamos (Fig. 1), portaban conspicuas manchas/sombras oscuras, especialmente en las celdillas marginales de las alas anteriores, incluso
alguna más tenue en las posteriores, estos elementos ya fueron citados por Monserrat (1982a, 1984g) sobre ejemplares de Cáceres y Almería, y es un hecho conocido en otras especies de este género tales como A. maculata Meinander, 1963, Aleuropteryx minuta Meinander, 1965, A. maculipennis Meinander, 1972 (Figs. 104, 112), A. vartianorum Aspöck & Aspöck, 1967 o A. argentata Tjeder, 1957. También McLachlan (1882b, 1882c) citaba ejemplares con alas manchadas de los Alpes, que podrían tratarse de la especie que ahora estamos estudiando, y Ward (1965) cita semejantes sombras en las alas anteriores de esta especie en ejemplares británicos, Aspöck & Aspöck (1999) y Zimmermann (2005b) en ejemplares austriacos, y Zelený (1964), Kis (1964a) o Meinander (1972a) lo mencionan para ésta y otras especies europeas, americanas y africanas. En cualquier caso, en lo que respecta a nuestra
fauna, en A. maculata y A. minuta (Figs. 85, 104, 112) (también en la especie canaria A. longiscapes Meinander, 1965) las manchas están bien definidas y aisladas, mientras que lo mencionado en ejemplares de A. juniperi, las manchas, a modo de sombras, son mucho más difusas y extendidas (Fig. 1).
Los estadios preimaginales de esta especie fueron descritos por Macleod (1964) como Helicoconis lutea, y posteriormente por Ward (1970: 75) y Henry (1976: 199).
NUEVO MATERIAL Y MATERIAL COMPLEMENTARIO ESTUDIADO: ESPAÑA: Almería, Gérgal, 30SWG4207, 860 m, 29.VI.2011, 1♂ a la luz, El Molino de La Serena, Bédar, 15.VII.2007, 1♂ sobre Rhamnus lycioides, 1♂ a la luz. Badajoz, Mérida, 29.IV.2006, 1♂ sobre Rosa sp., volando al amanecer. Cádiz, Facinas, 20.VI.2012, 2♂♂ a la luz, Pinsapar de Grazalema, 26.V.2012, 1♂ sobre Abies pinsapo. Ciudad Real, Navas de Estena, 8.VI.2012, 1♀ sobre Quercus faginea, F. Acevedo. Granada, Hoya de la Mora, 30SVG6506, 2500 m, 30.VI.2011, 2♂♂ sobre Pinus sylvestris, La Herradura, Cerro Gordo, 36°44’03.20’’N 3°46’03, 200 m, 1.VIII.2010, 1♀ sobre Pistacea lentiscus. Huelva, Matalascañas, 20.VI.2012, 1♂ sobre Juniperus phoenicea. Teruel, Laguna de Bezas, 6.VII.2010, 1♂ sobre J. thurifera, F. Acevedo (ver otro material en Helicoconis bachi n. sp.). ITALIA: Cerdeña, Sassari, Alghero, Capo Caccia, Dragunara, 40°34´32´´N, 8°09´39´´E, 10 m, 16.V.2008, 5♂♂, 9♀♀ sobre J. phoenicea, Fertilia, Monte Doglia, 40°36´37´´N, 8°14´43´´E, 190 m, 16.V.2008, 1♀ sobre Cupressus sempervirens, Sorso, Platamona, 40°49´10´´N, 8°31´02´´E, 10 m, 22.V.2008, 1♂, 1♀ sobre J. oxycedrus. A esta especie podría pertenecer una larva recolectada en Granada, Sierra Nevada, Hoya de la Mora (2550 m) en trampa de caída colocada bajo enebros rastreros (Juniperus communis) entre el 5.VII-1.IX.2010.
Aleuropteryx loewii Klapálek, 1894 (Figs. 31-35, 38, 125-128, 135-136)
Aleuropteryx löwii Klapálek, 1894
Klapálek, 1894: 121 (d).
Aleuropteryx loewii Klapálek, 1894
Klapálek, 1909: 30 (dis); Berland, 1962: 27 (rf); Aspöck & Aspöck, 1969: 40 (rf); Meinander, 1972a: 43 (rf); Monserrat, 1977a: 22 (list), 154 (key, mf), 155 (bio, dis), 1978b: 180 (bio, dis); Morgan, 1980: 119 (dis); Aspöck et al., 1980: 140 (bio, mf, tx, rf), Monserrat, 1980: 190 (bio, dis), 1982a: 81 (bio, dis), 1984a: 146 (dis, tx), 1984b: 104 (bio, dis), 1984c: 160 (bio, dis), 1984d: 41 (bio,
dis), 1984g: 26 (bio, dis), 1985b: 88 (bio, dis), 1985d: 128 (bio, dis), 1987a: 141 (bio, dis); Monserrat & Díaz-Aranda, 1987: 182 (bio, dis), 1988a: 92 (bio, dis); Díaz-Aranda & Monserrat, 1988a: 120 (bio, dis), 1988b: 225 (bio, dis), 1988c: 223 (bio, dis); Monserrat & Díaz-Aranda, 1989: 53 (bio, dis), 1990: 40 (bio, dis); Marín & Monserrat, 1990: 227 (bio, dis), 1991b: 182 (bio, dis); Monserrat & Marín, 1992: 282 (bio); Marín, 1994: 260-297 (bio, dis, fen); Monserrat et al., 1994: 55 (bio, dis); Marín & Monserrat, 1995a: 43 (bio, dis), 1995b: 118 (bio, dis); Monserrat, 1995: 110 (bio, dis), 1996a: 14 (list); H. Aspöck et al., 2001: 167 (rf); Monserrat, 2002: 4 (bio, dis), 2011: 165, 166 (bio, dis, tx), 2013: 299 (rf); Monserrat et al., 2013: 53 (bio, dis); Monserrat & Triviño, 2013: 20 (list), 116 (dis); Letardi et al., 2013: 36 (list).
Aleuropteryx loevi Klapálek, 1894 (lapsus)
Navás, 1921b: 65 (dis).
Aleuropteryx loewëi Klapálek, 1894 (lapsus)
Stitz, 1927: 17 (rf).
Aleuropteryx loewei Klapálek, 1894 (lapsus)
Eglin, 1940: 267 (rf).
Aleuropteryx loewi Klapálek, 1894
Navás, 1910a: 50 (rf), 1913a: 85 (dis), 1914c: 41 (dis), 1917b: 88 (dis), 1919a: 202 (dis), 1924a: 251 (dis, key, mf), 1925a:
127, 128 (dis, key, mf); Zelený, 1961a: 177 (rf); Meinander, 1963: 93 (rf); Ohm, 1973b: 303 (rf); Monserrat, 1977b: 368 (bio, dis), 1978a: 67 (bio, dis), 1979a: 416 (bio, dis); Díaz-Aranda et al., 1986a: 1125 (bio, dis), 1986b: 1141 (bio, dis); Monserrat & Triviño, 2013: 20 (list).
Aleuropteryx aequalis Navás, 1918
Navás, 1917b: 88 (dis, rf), 1918a: 233 (dis, rf); Anónimo (probablemente Navás), 1918: 172 (dis, rf), Navás, 1918c: 42 (d), 1919b: 43 (dis); Lacroix, 1924: 55 (rf); Tjeder, 1957: 111 (nt); Meinander, 1963: 93 (nt), 1972a: 43 (rf, tx); Aspöck et al., 2001: 166 (tx); Monserrat, 2011: 165 (tx); Monserrat & Triviño, 2013: 20 (list).
Aleuropteryx codinai Navás, 1910
Navás, 1910a: 49 (d), 54 (dis), 1921a: 152 (dis), 1921b: 65 (dis), 1924a: 251, 252 (dis, key, mf), 1925a: 127, 128 (dis, key,
mf), 1927b: 99 (dis); Lacroix, 1924: 55 (rf); Tjeder, 1957: 111 (rf); Meinander, 1963: 93 (nt); Meinander, 1972a: 39 (dis, list, nt, rf); Monserrat, 1977a: 158 (rf), 1977b: 372 (list, nt); Rausch & Aspöck, 1978a: 12 (rf); Aspöck et al., 1980: 9 (dis), 149 (mf, rf); Monserrat, 1984a: 150 (tx); Meinander, 1990: 3 (rf); Aspöck et al., 2001: 166 (tx); Monserrat, 2011: 164 (tx), 165 (mf); Monserrat & Triviño, 2013: 20 (list).
Elemento expansivo nor-mediterráneo. Está mayoritariamente citada sobre coníferas, especialmente del género Pinus, entre III-VIII, en altitudes de 10 hasta 1800 m. En nuestra fauna ha sido especialmente citada, incluyendo Baleares (Menorca),
sobre Pinus, también sobre Juniperus y Cedrus, en ocasiones sobre Ephedra, Arbutus, Quercus, Ceratonia, Ilex, Nerium, Pteridium, entre III-IX, en altitudes que oscilan entre 10-1900 m. Con frecuencia es extremadamente abundante.
Los estadios preimaginales de esta especie (Figs. 31-35, 38) fueron descritos por Löw (1885: 74), Lacroix (1924: 82), Rousset (1966: 71, 1969: 112) y Henry, 1976: 199.
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Almería, Albuñol, 30SVF8267, 70 m, 28.VI.2011, 2♂♂ sobre Pinus halepensis. Cádiz, Grazalema, 31.V.2005, 770 m, 7 ej., A. Gruppe (AG). Islas Baleares, Menorca, Cala Turqueta, 23.VII.2008, 1♂, 1♀ sobre P. halepensis, F. Acevedo. Granada, Aldeire, 30SVG9410, 1661 m, 8.V.2011, 1♂ sobre P. halepensis, Calahonda, 30SVF66, 20 m, 8.VII.2013, 1♂, ♀♀ sobre P. halepensis, Cáñar, 18.VII.2011, 1♂, 1♀ sobre P. sylvertris, Collado de la Sabina, 30SVG6207, 2050 m, 10.VII.2012, 1♂, 1♀ sobre P. sylvertris, Gualchos, 30SVF6462, 30 m, 28.VI.2011, 3♂♂ sobre P. halepensis, La Herradura, Cerro Gordo, 6.VIII.2011, 1♀ a la luz, 17.VIII.2013, 1♀ sobre P. halepensis, Los Tablones, 30SVF6481, 464 m, 6.V.2011, 1♂ sobre P. halepensis. Huelva, Matalascañas, 20.VI.2012, 2♂♂, 2♀♀ sobre P. pinea. Madrid, El Escorial, Abantos, 29.VI.2013, 1♂ sobre P. sylvestris, 19.VI.2014, 2♂♂, 2♀♀ sobre P. sylvestris, F. Acevedo. PORTUGAL: Beira Interior, Guarda, Sra. de Pestero, 800 m, 20.VI.2011, 1 ej., A. Gruppe (AG). Trásos-Montes e Alto Douro, Vila Real, Sa. Do Marão, 840 m, 18.VI.2011, 7 ej., A. Gruppe (AG).
Aleuropteryx maculata Meinander, 1963 (Figs. 112-120)
Meinander, 1963: 93 (d); Monserrat, 1984g: 26 (bio, dis, mf), 1985d: 128 (bio, dis), 1985e: 75 (rf); Díaz-Aranda et al., 1986a: 1125 (bio, dis), 1986b: 1141 (bio, dis); Monserrat & Díaz-Aranda, 1987: 182 (bio, dis); Díaz-Aranda & Monserrat, 1988b: 225 (dis); Meinander, 1990: 9 (rf); Marín & Monserrat, 1991b: 182 (bio, dis); Monserrat et al., 1991: 107 (rf); Monserrat & Marín, 1992: 282 (bio); Marín, 1994: 260-297 (bio, dis, fen); Marín & Monserrat, 1995a: 43 (bio, dis), 1995b: 118 (bio, dis); Monserrat, 1995: 110 (bio, dis), 1996a: 14 (list); Blasco, 2000: 98 (bio, dis); H. Aspöck et al., 2001: 169 (rf); Sziráki, 2004: 6 (rf); Monserrat & Triviño, 2013: 20 (list), 116 (dis).
Especie conocida de Marruecos y del centro, este y sur de España. Está mayoritariamente citada sobre Juniperus, Tetraclinis, a la luz y en trampa de Malaise, en II, V, VIII, en altitudes de hasta 1810 m. En nuestra fauna acompaña en su distribución
a enebros y sabinas, y ha sido citada sobre Juniperus communis, J. phoenica, en líquen sobre J. thurifera y también sobre esta conífera, entre V-IX (por muestreos seriados en un sabinar de Zaragoza, Monserrat, 1995 sugiere que algunas poblaciones puedan ser monovoltinas), en altitudes que oscilan entre 390-1376 m.
Que sepamos, los estadios preimaginales son desconocidos.
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Zaragoza, Retuerta de Pina, Pina de Ebro, 30TYL29, 400 m, 20.V.1991, 1♂ sobre Carduus bourgeanus Boiss. et Reut., J. Blasco.
Aleuropteryx minuta Meinander, 1965 (Figs. 104-111)
Meinander, 1965b: 56 (d); Monserrat, 1984g: 26 (rf), 1985d: 127 (rf), 128 (bio, dis, mf), 1985e: 75 (rf), 1987a: 141 (bio, dis), 145 (rf); Meinander, 1990: 9 (rf); Monserrat & Díaz-Aranda, 1990: 40 (bio, dis); Monserrat et al., 1991: 108 (rf); Monserrat & Marín, 1992: 282 (bio); Monserrat, 1995: 111 (bio, dis); H. Aspöck et al., 2001: 168 (rf); Monserrat, 2002: 4 (bio, dis), Sziráki, 2004: 6 (rf) ; Monserrat, 2010: 26 (bio, dis), 2013: 300 (rf); Monserrat & Triviño, 2013: 20 (list), 117 (dis).
Especie conocida de la República Árabe Saharaui, Marruecos y del S. E. de España (Almería), fuertemente asociada con zonas
térmicas, secas y hábitats hialinos, ramblas y zonas de malpaís. Especialmente recolectada a la luz y sobre Tamarix, también ha sido citada sobre Phoenix, Zizyphus, Pistacea y Ceratonia, entre V-VIII, en altitudes de entre 10 m hasta 1330 m. En nuestra fauna ha sido citada en similares ambientes (Almería)
sobre Tamarix, entre IV-VIII (probablemente bivoltina), en altitudes que oscilan entre 40-555 m.
Que sepamos, los estadios preimaginales son desconocidos.
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Almería, Casillas de Atochares, 30SWF79, 206 m, 6.VII.2013, 1♀ sobre Tamarix sp., Tabernas, Rambla Roja, 30SWG50, 400 m, 13.VI.2012, 1♂ a la luz, F. Acevedo.
Aleuropteryx remane Rausch, Aspöck & Ohm, 1978 (Figs. 129-132, 137-138)
Rausch et al., 1978b: 45 (d); Aspöck et al., 1980: 9 (dis), 141 (bio, mf, tx, rf); Monserrat, 1984b: 113 (rf), 1985b: 92 (rf), 1987a: 141 (bio, dis), 145 (rf); Meinander, 1990: 8 (rf); Monserrat & Díaz-Aranda, 1990: 41 (bio, dis); Monserrat & Marín, 1992: 282 (bio); Monserrat et al., 1994: 56 (bio, dis); Marín & Monserrat, 1995b: 118 (bio, dis); Monserrat, 1995: 112 (bio, dis), 1996a: 14 (list); H. Aspöck et al., 2001: 168 (rf); Monserrat, 2002: 5 (bio, dis); Sziráki, 2004: 5 (rf); Monserrat, 2013: 300 (rf); Monserrat et al., 2013: 53 (bio, dis); Monserrat & Triviño, 2013: 20 (list), 117 (dis).
Especie ibérica conocida del sur-este peninsular (Murcia, Almería, Granada, Alicante) y de otras zonas más septentrionales
de la Peninsula Ibérica (Madrid, Zaragoza, Navarra, Lérida), poco citada y generalmente recolectada a la luz en zonas secas
y térmicas carentes de vegetación arbórea. Sorprende su versatilidad al haber sido hallada tanto en zonas xéricas, como marcadamente
pirenaicas. A veces citada sobre Artemisia herba-alta, Quercus coccifera y Q. faginea, o en sabinares, ocasionalmente emergiendo de agallas de Ptiloedaspis tavaresiana Bezzi, 1920 (Diptera: Tephritidae), entre V-IX (probablemente sufra diapausa estival), en altitudes de 10-1310 m.
Que sepamos, los estadios preimaginales son desconocidos.
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Alicante, Olmeda del Prado, 560 m., 5.VIII.2008, 1 ej., Lencina (AG). Almería, El Molino de La Serena, Bédar, 17.VII.2007, 2♀♀ a la luz, Rambla del Retamar, 14.VI.2012, 1♀ a la luz, V. Triviño, Tabernas,
Rambla Roja, 30SWG50, 400 m, 13.VI.2012, 1♂, 4♀♀ a la luz, V. Triviño, 14.IX.2013, 1♂ a la luz, F. Acevedo. Murcia, Jumilla, Sa. del Carche, 990 m, 22.VII.2008, 1 ej., Lecina & Gallego (AG).
Helicoconis hirtinervis Tjeder, 1960 (Figs. 145, 146)
Tjeder, 1960: 313 (d); Monserrat & Triviño, 2013: 20 (list).
Helicoconis (Helicoconis) hirtinervis Tjeder, 1960
Monserrat et al., 1994: 56 (bio, dis); H. Aspöck et al., 2001: 171 (rf); Monserrat & Triviño, 2013: 20 (list), 128 (dis); Tillier, 2013: 141 (rf); Vidlička, 2014: 442 (rf); Tillier, 2015: 590 (rf).
Elemento conocido de Alemania, Dinamarca, Escocia, Suiza, Italia, Francia, España, Slovenia y Turquía. Está mayoritariamente
citada sobre Picea abies, también sobre Stipa, entre V-VIII, en altitudes montanas de 1680 m hasta 1992 m. En nuestra fauna ha sido únicamente citada en Lérida, sobre
avellano, en VII, a 1890 m de altitud.
Los estadios preimaginales de esta especie fueron “descritos” por Nielsen, 1979.
Helicoconis hispanica Ohm, 1965 (Figs. 147-149)
Ohm, 1965: 196 (d); Aspöck et al., 1980: 145 (bio, mf, tx, rf); Monserrat, 1984b: 106 (bio, dis), 1984d: 42 (bio, dis), 1985d: 131 (bio, dis), 1987a: 142 (bio, dis); Díaz-Aranda & Monserrat, 1988c: 224 (dis); Monserrat & Díaz-Aranda, 1990: 41 (bio, dis); Monserrat & Marín, 1992: 282 (bio); Lo Verde & Monserrat, 1997: 59 (rf); Nicoli Aldini et al., 2012: 448 (rf); Monserrat & Triviño, 2013: 20 (list).
Helicoconis (Fontenellea) hispanica Ohm, 1965
Meinander, 1972a: 129 (dis, mf, rf), 1990: 25 (rf); Marín & Monserrat, 1995b: 118 (bio, dis); Monserrat, 1995: 113 (dis), 1996a: 14 (list); H. Aspöck et al., 2001: 173 (rf); Sziráki, 2004: 18 (rf); Monserrat, 2013: 302 (bio, dis); Monserrat & Triviño, 2013: 20 (list), 127 (dis); Tillier, 2013: 141 (rf), 142 (bio, dis), 2015: 590 (rf).
Especie conocida de España, Francia, Italia (Cerdeña, Sicilia) y Marruecos, generalmente asociada a zonas térmicas y secas.
Escasamente citada y mayoritariamente recolectada a la luz, algunos especímenes lo han sido sobre Juniperus, Pinus, Crataegus, Quercus y Olea, aunque probablemente viva en vegetación baja o herbácea, y ha sido recolectada entre VI-VIII, en altitudes entre 370 hasta
1992 m. En nuestra fauna ha sido recolectada en el sur y levante, mayoritariamente a la luz, en un sabinar, o sobre Pinus, Juniperus, Quercus, Olea y Crataegus, entre VI-X (probablemente sus poblaciones tengan diapausa estival o simplemente sean monovoltinas), en altitudes que oscilan
entre 10-1750 m.
Que sepamos, los estadios preimaginales son desconocidos.
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Almería, San José, Playa de Mónsul, 28.VI.2011, 3♂♂ a la luz, 20.VII.2011, 2♂♂ a la luz, Tabernas, Rambla Roja, 30SWG50, 400 m, 13.VI.2012,
1♂ a la luz, F. Acevedo.
Helicoconis iberica Ohm, 1965 (Figs. 150, 151)
Ohm, 1965: 198 (d); Aspöck et al., 1980: 9 (dis), 146 (bio, mf, tx, rf); Díaz-Aranda & Monserrat, 1988c: 224 (rf); Monserrat & Marín, 1992: 282 (bio); Sziráki & Harten, 2006: 394 (rf); Monserrat, 2010: 28 (rf); Monserrat & Triviño, 2013: 20 (list).
Helicoconis (Fontenellea) iberica Ohm, 1965
Meinander, 1972a: 131(dis, mf, rf), 1977: 81, 84 (rf), 1990: 25 (rf); H. Aspöck et al., 2001: 173 (rf); Sziráki, 2004: 18 (rf); Sziráki & Harten, 2006: 394 (rf); Monserrat, 2013: 303 (rf); Monserrat & Triviño, 2013: 20 (list), 127 (dis).
Elemento muy poco conocido, citado de la Península Ibérica y de Yemen. Está mayoritariamente recolectada a la luz, también
sobre Juniperus, entre III-VII, en altitudes de entre 1200-1300 m. En nuestra fauna ha sido muy poco citada (Sa. de Alfacar en Granada), también mayoritariamente recolectada a la luz, en VII, en altitudes que oscilan entre 1200-1250 m.
Que sepamos, los estadios preimaginales son desconocidos.
Helicoconis interna (Navás, 1911) (Figs. 152-154)
Aleuropteryx interna Navás, 1911
Navás, 1911: 209 (d), 1913a: 86 (tx); Lacroix, 1924: 55 (rf); Enderlein, 1930: 114 (rf); Tjeder, 1957: 111 (rf); Meinander, 1963: 92 (nt); Ohm, 1965: 171 (rf), 205 (nt, tx); Monserrat & Triviño, 2013: 20 (list).
Helicoconis interna (Navás, 1911)
Navás, 1913a: 86 (tx), 1924a: 249 (dis, key, mf), 1925a: 126 (dis, key, mf); Kis, 1965a: 125 (nt, rf); Ohm, 1965: 171 (rf), 205 (list, nt, rf); Meinander, 1972a: 116 (nt, rf); Monserrat, 1977c: 139 (nt); Meinander, 1990: 3 (rf), 23 (list); Monserrat, 2011: 166 (dis, tx).
Helicoconis (Ohmopteryx) interna (Navás, 1911)
Monserrat, 2013: 302 (bio, dis); Monserrat et al., 2013: 54 (bio, dis); Monserrat & Triviño, 2013: 20 (list), 129 (dis).
Helicoconis laufferina Navás, 1913
Navás, 1913a: 85 (d); Lacroix, 1924: 55 (rf); Navás, 1924a: 249 (dis, key mf), 1925a: 126, 127 (dis, key, mf), 1929b: 125 (dis); Enderlein, 1930: 114 (nt, tx); Tjeder, 1957: 106 (rf, tx), 1960: 312 (tx); Meinander, 1963: 92 (nt); Kis, 1965a: 125 (nt, rf), 129 (rf); Ohm, 1965: 171 (rf), 205 (list, nt, tx); Meinander, 1972a: 116 (nt, rf); Monserrat, 1977a: 22 (list), 152 (nt), 1977c: 139 (nt), 1984d: 47 (nt); Meinander, 1990: 3 (rf), 23 (list, rf); Monserrat, 1996a: 16 (list, tx), 2011: 168 (tx); Monserrat & Triviño, 2013: 20 (list).
Helicoconis pseudolutea Ohm, 1965
Aspöck & Aspöck, 1964: 154 (rf); Ohm, 1965: 188 (d); Kis, 1965a: 129 (rf); Monserrat, 1977a: 22 (list), 152 (rf), 153 (bio, dis), 1978a: 68 (bio, dis), 1979a: 416 (bio, dis); Aspöck et al., 1980: 145 (bio, mf, tx, rf); Monserrat, 1982a: 82 (bio, dis), 1984b: 105 (bio, dis), 1984c: 160 (bio, dis), 1984d: 42 (bio, dis), 1984e: 175 (bio, dis), 1984g: 28 (bio,
dis), 1985b: 89 (bio, dis), 1985d: 130 (bio, dis); Díaz-Aranda et al., 1986a: 1125 (bio, dis), 1986b: 1142 (bio, dis); Monserrat, 1987a: 142 (bio, dis); Marín & Monserrat, 1987: 350-359 (bio, dis); Monserrat & Díaz-Aranda, 1987: 182 (bio, dis), 1988a: 93 (bio, dis); Séméria & Berland, 1988: 50 (rf); Díaz-Aranda & Monserrat, 1988a: 120 (bio, dis), 1988b: 226 (bio, dis), 1988c: 223 (bio, dis); Monserrat & Díaz-Aranda, 1989: 53 (bio, dis), 1990: 41(bio, dis); Marín & Monserrat, 1990: 227 (bio, dis), 1991b: 182 (dis); Monserrat & Marín, 1992: 282 (bio); Marín, 1994: 260-297 (bio, dis, fen); Monserrat, 2010: 28 (rf), 2011: 167 (tx); Monserrat & Triviño, 2013: 20 (list); Letardi et al., 2013: 36 (list).
Helicoconis (Ohmopteryx) pseudolutea Ohm, 1965
Kis et al., 1970: 64 (rf); Meinander, 1972a: 128 (rf); Monserrat, 1978b: 182 (bio, dis), 1980: 191 (bio, dis), 1995: 113 (bio, dis); Marín & Monserrat, 1995b: 118 (bio, dis); Monserrat, 1996a: 14 (list); H. Aspöck et al., 2001: 172 (rf); Monserrat, 2002: 5 (bio, dis); Monserrat & Triviño, 2013: 20 (list).
Especie conocida del S.O. de la Región Paleártica (centro y sur de Europa, Marruecos, Túnez, Anatolia, Cáucaso, Irak e Irán),
generalmente asociada a zonas térmicas y xéricas. Mayoritariamente recolectada a la luz, aunque también sobre Juniperus, Larix, Pinus, Populus, Castanea, Quercus, Ceratonia, Daphne, Ulmus, Celtis, Corylus, Berberis, Crataegus, Retama,
Betula, Pistacea y Olea, aunque probablemente viva en vegetación herbácea, entre III-VIII, en altitudes de 20 hasta 2200 m. En nuestra fauna ha sido
citada sobre estas plantas entre V-IX (a pesar de esta amplitud, Monserrat, 1995, por los datos obtenidos en seguimientos poblacionales mediante trampas de Malaise, sugiere ciclos monovoltinos), en altitudes
que oscilan entre 130-1900 m.
Merecen citarse en algunos ejemplares ahora citados (tanto ♀ como ♂) de España, Madrid (Chinchón) la presencia de una venilla transversal entre Mp + CUa-CUp en las alas posteriores, elemento nada usual en este
género (Fig. 86) y característico, entre los de nuestra fauna de esta subfamilia, del género Aleuropteryx (Figs. 23, 85).
Que sepamos, los estadios preimaginales son desconocidos.
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Almería, Turrillas, 13.VI.2012, 1♂, 1♀ sobre Quercus rotundifolia, F. Acevedo. Badajoz: Valverde de Burguillos, 23.V.2012, 1♂, 2♀♀ sobre Q. rotundifolia. Cádiz, Algodonales, 26.V.2012, 1♂ sobre Q. rotundifolia, El Bosque, 21.VI.2012, 1♀ a la luz, Los Alcornocales, 21.VI.2012, 1♂, 1♀ sobre Q. suber, Pinsapar de Grazalema, 22.VI.2012, 2♂♂ sobre Abies pinsapo, 1♀ sobre Q. rotundifolia, Puerto del Boyar, 22.VI.2012, 1♀ sobre Q. rotundifolia, Ubrique, 25.V.2012, 2♂♂ sobre Q. canariensis. Granada, Cáñar, 30SVF6389, 1354 m, 27.VI.2011, 1♂ sobre Q. pyrenaica, 18.VII.2011, 2♂♂ sobre Q. pyrenaica, El Dornajo, 30SVG60, 1750 m, 10.VII.2012, 2♂♂ sobre Q. rotundifolia, 10.VII.2013, 2♂♂, 2♀♀ sobre Q. rotundifolia. Huelva: La Corte, 23.V.2012, 1♂ sobre Q. rotundifolia, La Nava, 23.V.2012, 3♀♀ sobre Q. rotundifolia, Patrás, 19.VI.2012, 1♀ sobre Eucaliptus sp. Madrid, Chinchón, El Salitral, 40° 10’ 73’’N 3° 28′76’’ O, 560 m, 6.VI.2009, 1♀ volando al atardecer, 20.VI.2009, 2♂♂ a la luz,
Hoyo de Manzanares, 14.VI.2014, 1♂ sobre Q. ilex, F. Acevedo, 1♂ sobre Juniperus oxycedrus, F. Acevedo. Málaga, Las Asomadillas, 25.V.2012, 1♂ sobre Pistacea lentiscus, Puerto de Montejaque, 22.VI.2012, 1♀ sobre Q. rotundifolia.
Helicoconis panticosa Ohm, 1965 (Figs. 155-157)
Ohm, 1965: 200 (d); Monserrat, 1979a: 416 (bio, dis); Aspöck et al., 1980: 146 (bio, mf, tx, rf); Monserrat, 1984b: 105 (bio, dis), 1984c: 160 (bio, dis), 1984d: 47 (rf), 1984g: 28 (bio, dis), 1985b: 88 (bio, dis); Monserrat & Marín, 1992: 282 (bio); Ribera & Blasco-Zumeta, 1998: 975 (bio, dis), 978 (nt); Monserrat & Triviño, 2013: 20 (list).
Helicoconis (Fontenellea) panticosa Ohm, 1965
Meinander, 1972a: 133 (dis, mf, rf), 1990: 25 (rf); Monserrat et al., 1994: 56 (bio, dis); Marín & Monserrat, 1995a: 43 (bio, dis); Monserrat, 1995: 113 (dis), 1996a: 14 (list); H. Aspöck et al., 2001: 173 (rf); Sziráki, 2004: 19 (rf); Monserrat & Triviño, 2013: 20 (list), 128 (dis).
Elemento ibérico extendido hasta Turquía. Está mayoritariamente citada a la luz, entre VI-VII, en altitudes de entre 750-2000
m. En nuestra fauna ha sido muy escasamente citada, ha sido recolectada sobre alcornoques, robles, pinsapos y pinos, entre
V-IX, en altitudes que oscilan entre 10-2000 m. Sorprende de esta especie su versatilidad, ya que la encontramos desde medios
alpinos (Panticosa, 2000 m, Bosost, 1150 m) a medios subdesérticos o mediterráneos extremadamente térmicos (Los Monegros,
200 m, Pilar de la Horadada, 10 m).
Que sepamos, los estadios preimaginales son desconocidos.
Helicoconis bachi n. sp. (Figs. 139-144, 158)
http://urn:lsid:zoobank.org:act:0CE47E93-FFD8-468B-87E4-393BE2556F25
Aleuropteryx sp.
Janetschek, 1957: 148-149 (dis, mf); Meinander, 1990: 9 (rf); Monserrat, 2011: 166 (rf), 2013: 304 (rf); Monserrat & Triviño, 2013: 11 (rf), 20 (list).
Helicoconis sp.
Monserrat & Triviño, 2013: 11 (rf), 20 (list), 129 (dis).
MATERIAL ESTUDIADO: Holotipo: ESPAÑA: Granada, Sierra Nevada, Observatorio Viejo del Veleta, 37°05’26.94’’N 3°22’55.06’’O, 2580 m, 1♂ en trampa de caída con líquido de
Turquin colocada entre el 29.VI-25.VIII.2011 bajo enebros rastreros (Juniperus communis). En la colección del autor, depositada en la Facultad de Biología de la Universidad Complutense (Madrid), con el número
de referencia CO-3612.
Paratipos: Estén donde estén los dos ejemplares citados por Janetschek (1957), probablemente un macho y una hembra, que aún no han sido localizados, los consideramos paratipos de la especie que ahora
describimos.
DIAGNOSIS:
Macho: Longitud de las alas anteriores: 3,5 mm, de las posteriores: 2,7 mm. Genitalia: ectoprocto con procesos ventrales muy desarrollados,
digitiformes y levemente recurvados hacia la línea media (Figs. 139, 140). Estilos muy cortos y robustos, bifurcados con extremos cortos, trangulares y levemente recurvados hacia arriba y hacia
la línea media (Figs. 139-142, 158). Hipandrio con margen caudal levemente convexo en su zona media (Fig. 140). Pene más largo que los parámeros, en vista lateral, adelgazado hacia su extremo caudal de forma asimétrica respecto a su
eje, y con una globosa formación membranosa ventral (Figs. 141, 143). Parámeros espatulados y levemente arqueados (Figs. 141, 144).
Hembra: Desconocida, presuntamente braquípteras.
DESCRIPCIÓN:
Machos: Cabeza pardo oscura con región frontal e interantenal escasamente esclerificada. Ojos negros, de tamaño habitual. Palpos
labiales pardos oscuros, palpos maxilares pardos, más pálidos los dos últimos segmentos. Antenas con escapo pardo, fusiforme,
algo mayor que el pedicelo, éste también pardo y rectangular. Flagelo con 25 flagelómeros, más oscuros y más anchos que largos
los más basales y progresivamente van tornando a más pálidos y más largos que anchos, el último alargado y ovoide en su extremo.
Flagelómeros 11°-12° y 14°-15° en una de las antenas, y 16°-17° y 19°-20° en la otra, algo más cortos, ofreciendo aspecto
de ser flagelómeros duplicados.
Tórax pardo rojizo oscuro. Patas con coxas y trocánteres pardos oscuros, resto muy pálidas, casi hialinas, algo más largas
que lo habitual observado en otras especies. Alas hialinas, sin apenas desarrolladas las setas marginales. Longitud de las
alas anteriores: 3,5 mm, de las posteriores: 2,7 mm.
Abdomen pardo muy pálido. Genitalia con ectoprocto portando abundante setación y con procesos ventrales muy desarrollados,
digitiformes y levemente recurvados hacia la línea media (Figs. 139, 140). Estilos muy cortos y muy robustos, bifurcados apicalmente con extremos cortos, trangulares y levemente recurvados hacia
arriba y hacia la línea media (Figs. 139-142, 158). Hipandrio con margen caudal levemente convexo en su zona media (Fig. 140) y proceso ventral del IX segmento digitiforme y bien desarrollado (Figs. 139, 140). El pene, bastante más largo que los parámeros, en vista lateral, está adelgazado hacia su extremo caudal de forma asimétrica
respecto a su eje, y porta una globosa formación membranosa ventral (Figs. 141, 143). Parámeros espatulados y levemente arqueados, con extremo caudal acodado hacia afuera (Figs. 141, 144).
Hembras: Desconocidas, presuntamente braquípteras.
Estadios larvarios: Desconocidos.
DERIVATIO NOMINIS: Dedico esta nueva especie a Johann Sebastian Bach (1685-1750), por tantas cosas gracias a él disfrutadas y compartidas durante
toda mi vida..., pero sobre todo por hacerme escuchar muchas veces, como hombre, la voz de quien, como científico, no creo.
DISCUSIÓN: Antes de pasar a la discusión propiamente dicha de esta nueva especie, consideramos necesario comentar el largo camino de
pesquisas y esfuerzos que han sido necesarios para conocer la identidad de esta recóndita y esquiva especie.
Todo parece comenzar cuando en el verano (15.VII-4.VIII) de 1954 un grupo de investigadores austríacos realizaron una serie
de muestreos en Sierra Nevada (Granada) a resultas de los cuales, y poco después, Janetschek (1957) mencionaba de la ladera norte del Veleta (c. 2400-2720 m) dos pequeños ejemplares, uno de ellos braquíptero, de una probable
“nueva especie” de Coniopterygidae que fue citada como Aleuropteryx spec. Que sepamos, el asunto permaneció en silencio hasta que fue mencionado por Meinander (1990) y Monserrat (2011) hace referencia a este dato.
Desde nuestro conocimiento sobre esta información, y prácticamente desde que iniciamos hace casi 40 años nuestros estudios
sobre los neurópteros de la Fauna Ibérica, hemos intentado recolectar nuevos ejemplares en Sierra Nevada (desde Díaz-Aranda
& Monserrat, 1988c) y zonas próximas para conseguir nuevos ejemplares sobre los que dilucidar esta enigmática especie. Parte de este enorme
esfuerzo es reflejado por Monserrat (2011, 2013) quien anota los muestreos realizados para tratar de obtener nuevos individuos, y sospecha que podrían tratarse de ejemplares
asignables al género Heteroconis, que en ocasiones ha sido citado con especies braquípteras o ápteras de este género en otras zonas elevadas de otros países
y continentes próximos (Europa, Canarias, Marruecos, Turquía, etc.) (ver más adelante), y no hemos cejado desde entonces en
este empeño, y especialmente durante los últimos 10 años, en los que año tras año y hasta la actualidad, hemos venido realizado
muestreos en esta zona utilizando diferentes tipos de trampas de caída con líquido de Turquin, tomando de muestras de diversos
tipos de hojarascas y bateando Pinus y especialmente de Juniperus existentes a esa elevada altitud, sin más resultados, hasta hoy, que un único ejemplar.
Paralelamente, y desde hace varios años, hemos venido haciendo gestiones para tratar de localizar este material original,
especialmente con la consulta a diversos colegas austríacos que nos pudieran proporcionar algún dato sobre el paradero de
estos ejemplares, y a diferentes centros de investigación de este país, donde en sus colecciones pudieran haber permanecido.
Heinz Janetschek (1913-1997) fue profesor en la universidad de Innsbruck (Thaler, 1997), por lo que centramos nuestras pesquisas en esta universidad, ya que, de existir aún este material, podría haber permanecido
en las colecciones de este centro. Merced a la amabilidad y gestiones de Hubert y Renate Rausch del Naturkundliche Gesellschaft
Mostviertel (Scheibbs, Austria) pudimos conocer que Birgit Schlick-Steiner y Florian Steiner del Instituto de Ecología de
la Universidad de Innsbruck, les habían informado de que los dos ejemplares de Aleuropteryx (que Janetschek había publicado en 1957) no se hallaban en las colecciones de la citada universidad (aunque sí existía otro
material de aquella excursión a Sierra Nevada de 1954), y comentaban que tales ejemplares habían sido estudiados en su día
por D. E. Kimmins, y que quizás pudieran haber sido trasladados y permanecido en el Natural History Museum en Londres, donde
se conserva gran parte de su material como legado. Por ello me puse en contacto con Ben Price del Natural History Museum (Londres,
Reino Unido) quien amablemente me comentó que tampoco allí se habían localizado estos ejemplares. Estén donde estén estos
ejemplares, y a la espera de que puedan aparecer en el futuro, los consideramos paratipos de la especie que ahora describimos.
Sobre la discusión de esta especie, comentemos inicialmente que el braquipterismo, como adaptación a la altitud es, entre
otros condicionantes, un hecho sobradamente conocido entre los insectos (Mani, 1968), y aunque no siempre lo hallamos en especies de alta montaña, no es infrecuente entre los neurópteros, sea Dilaridae o en
particular Hemerobiidae (Oswald, 1993, 1996; Pantaleoni & Letardi, 1996; Tauber et al., 2007; Monserrat, 2015), y ha sido ocasionalmente citado en esta familia, bien característico de especies de algún género con reducción de las alas
posteriores (Conwentzia) o de ambas alas (Coniopteryx), y particularmente ha sido citado en el género que tratamos, en el que varias especies ápteras o braquípteras han sido descritas
o citadas de Europa (Bulgaria, Dinamarca), Continente Africano (Islas Canarias, Marruecos, Uganda), y Continente Asiático
(Turquía, Turkmenistán) etc. (Carpenter & Lestage, 1928; Kimmins, 1950; Tjeder, 1960; Messner, 1965; Ohm, 1965; Meinander, 1972a; Rausch et al., 1978a; Monserrat, 2002), característica que se da con mayor frecuencia e intensidad en las hembras, a veces también el macho, y en nuestro caso,
y a tenor de los datos ahora anotados y de la información de Janetschek (1957), es deducible que en Helicoconis bachi n. sp. sean braquípteras solo las hembras. En cualquier caso, el braquipterismo no es un elemento correlacionado con un tipo determinado
de estructuras genitales, por lo que hay que recurrir a ellas para su discusión.
Han sido descritas del género Helicoconis unas 35 especies, algunas transferidas posteriormente a otros géneros, y hoy día incluye unas 25 especies, de las que 16
habitan en el Paleártico occidental (Meinander, 1972a, 1990; Monserrat & Díaz-Aranda, 1988b; Aspöck et al., 2001; Monserrat, 2002; Sziráki, 2004; Oswald, 2013a; Vidlička, 2014), muchas de ellas aisladas y adaptadas a zonas de alta montaña, reflejo de distribuciones preglaciares más extensas y que
ha generado numerosos procesos de especiación.
Excepción hecha de Helicoconis aptera Messner, 1965, conocida de Bulgaria, Turquía y Turkmenistán, cuya genitalia masculina es desconocida, y a pesar de lo poco consistentes
y en ocasiones compartidos entre unas y otras las características que “definen” los grupos de especies (subgéneros según algunos
autores), entre las especies restantes, la ausencia de proceso basal en el estilo de la genitalia masculina de Helicoconis bachi n. sp. (Figs. 140, 141) la aleja de las especies que sí lo poseen (grupo Helicoconis s.str. autores) (Fig. 145), y la ausencia de escotadura y dentículo dorsales en los parámeros (Fig. 143) también la diferencia de las especies que sí los poseen (grupo Ohmopteryx autores) (Fig. 153), siendo pues más próxima a las restantes nueve especies (grupo Fontenellea autores), conocidas desde las Islas Canarias (Tenerife), norte y centro de África (Marruecos, Argelia, Uganda), sur de Europa
(Península Ibérica, Italia, Hungría, Rumanía) y Asia occidental y central (Anatolia, Arabia Saudí, Yemen, Mongolia). Entre
ellas, quedan claramente diferenciadas y descartadas las especies con estilos no o mínimamente bifurcados en su extremo (Figs. 147, 151), acercándola pues a las especies que poseen los estilos marcadamente bifurcados en su extremo (Figs. 152, 157). Con estas características, conocemos cuatro especies:
Helicoconis sengonca Rausch, Aspöck & Aspöck, 1978, conocida de Anatolia, que posee estilos con ramas caudales muy largas, parámeros bifurcados
y curvos en su alargado extremo caudal, pene muy delgado y alargado en su extremo caudal, y margen caudal del hipandrio dentado,
elementos que no aparecen en Helicoconis bachi n. sp. (Figs. 140, 141, 143, 144, 158).
Helicoconis canariensis Monserrat, 2002, conocida de Tenerife, en la que tanto el macho como la hembra son braquípteros, y en el macho sus estilos son bastante más
largos, con diente interno romo, no agudo, pene acodado en su extremo caudal, no agudizado, extremo distal largo y delgado,
no corto y cónico, sin procesos membranosos ventrales, y parámeros con diente ventral, elementos diferentes a los que aparecen
en Helicoconis bachi n. sp. (Figs. 140, 141, 143, 144, 158).
Más similar parece Helicoconis algirica Meinander, 1976, descrita (muy someramente) de Argelia, en la que el hipandrio carece de proyección caudal, el extremo caudal de los parámeros
está fuertemente acodado y el del pene es romo, elementos que no aparecen en Helicoconis bachi n. sp. (Figs. 140, 141, 143, 144, 158).
Por último mencionemos Helicoconis panticosa Ohm, 1965, descrita y conocida de la Península Ibérica (desde Pirineos al sureste peninsular) y citada de Anatolia, que parece ser
la especie más próxima. En la genitalia masculina de esta especie los estilos son mucho más largos y estilizados (Fig. 157), los parámeros son rectos y estrechos (Fig. 155), el pene, en vista lateral, está adelgazado hacia su extremo caudal de forma casi simétrica respecto a su eje, sin portar
una patente globosa formación membranosa ventral (Fig. 156), mientras que en Helicoconis bachi n. sp. los estilos son mucho más más cortos y robustos (Figs. 140, 158), los parámeros son curvos y espatulados (Fig. 144), y el pene, en vista lateral, está adelgazado hacia su extremo caudal de forma asimétrica respecto a su eje, portando una
globosa formación membranosa ventral (Figs. 141, 143).
Estando este artículo en prensa, y tratando de recolectar algún nuevo ejemplar de Helicoconis bachi n. sp., prosiguieron los muestreos durante 2016. Así, en la vertiente norte del Veleta, Sierra Nevada, Granada (Mojón del Trigo, 30S04662274105, 2.551 m) se instalaron 11 trampas de caída con glicerina + cerveza, bajo enebros y sabinas,
que estuvieron instaladas desde el 27.VI.2016 hasta el 23.VII.2016, sin conseguir ningún nuevo ejemplar. Igualmente, en esta
localidad, el 27.VI.2016, y con aspirador portátil, se muestreó otra vegetación presente en la zona: sobre gramíneas, Hormathophylla spinosa (Brassicaceae), Erygium glaciale (Apiaceae), Heliamthemum sp. (Cistaceae), sin ningún resultado. No así sobre Juniperus sabina y Juniperus communis (Cupressaceae), que bien con este método o bien mediante batido con manga entomológica, se obtuvieron 3 ♀♀ de Semidalis pseudouncinata Meinander, 1963 y 6 ♀♀ y 2 ♂♂ de Aleuropteryx juniperi Ohm, 1968 (en esta especie y en ambos sexos con membrana de las alas oscura y con manchas difusas, especialmente marcadas en las celdillas
marginales de las anteriores). Esta operación (tanto mediante aspirador, como de batido con manga entomológica sobre enebros
y sabinas) se repitió el 23.VII.2016, obteniéndose 2 ♂♂ y 5 ♀♀ de Aleuropteryx juniperi Ohm, 1968 (con similar tipo de manchas oscuras). En cualquier caso no se consiguió, una vez más, el objetivo de hallar nuevos ejemplares
para incrementar la serie tipo de esta esquiva especie.
Es cierto que nos hubiera gustado contar con un mayor número de ejemplares, pero no se puede dudar de que el esfuerzo realizado
para obtenerlos ha sido titánico. Quizás no hemos acertado con la planta/s sobre las que pueden desarrollarse, pero no son
demasiadas (ni fáciles de muestrear) las existentes a estas altitudes (2580 m) en Sierra Nevada, y “a nuestro favor” citemos
que la mayoria de las especies conocidas de este género, y no solo las de alta montaña, han sido descritas a partir de un
único ejemplar, o solo se conocen de un único macho, hechos que sugieren una baja tasa de elementos en sus poblaciones y sin
duda la dificultad de recolectarlos en estos montanos medios.
Coniopteryginae
Coniopteryx (Coniopteryx) ezequi Monserrat, 1984 (Figs. 24, 159-166)
Monserrat, 1984a: 147 (dis, tx), 1984g: 29 (d), 1985c: 240 (list); Díaz-Aranda et al., 1986a: 1125 (bio, dis), 1986b: 1142 (bio, dis); Monserrat & Díaz-Aranda, 1987: 183 (bio, dis), 1988a: 93 (bio, dis); Díaz-Aranda & Monserrat, 1988b: 226 (bio, dis); Monserrat & Díaz-Aranda, 1989: 54 (bio, dis), 1990: 43 (bio, dis); Meinander, 1990: 55 (rf); Marín & Monserrat, 1990: 227 (bio, dis), 1991b: 183 (bio, dis); Marín & Monserrat, 1995a: 44 (bio, dis), 1995b: 119 (bio, dis); Monserrat, 1995: 120 (bio, dis), 1996a: 15 (list); H. Aspöck et al., 2001: 184 (rf); Monserrat, 2002: 12 (bio, dis); Monserrat & Triviño, 2013: 20 (list), 118 (dis).
Coniopteryx ezequi Monserrat, 1984
Monserrat & Marín, 1992: 283 (bio); Marín, 1994: 260-297 (bio, dis, fen); Blasco, 2000: 98 (bio, dis); Sziráki, 2004: 30 (rf); Monserrat & Triviño, 2013: 20 (list).
Especie conocida de la mitad oriental de la Península Ibérica y citada erróneamente de Guinea Ecuatorial (Isla de Bioko) por
Monserrat (1995) que sin duda se trata de una confusión de etiquetación de ejemplares (Monserrat, 2002). Por primera vez se cita de zonas meridionales de la Península Ibérica, lo que hace presumir una mayor extensión en ella.
Fuertemente asociada con enebros y sabinas (Juniperus) a los que acompaña en su distribución, y ocasionalmente recolectada sobre Pinus, Tamarix y a la luz. Parece una especie muy estenotópica y local, pero a veces es muy abundante y fácilmente visible al atardecer
volando alrededor de enebros y sabinas. Los datos ahora aportados mantienen estas características. Está mayoritariamente citada
entre III-IX (Monserrat, 1995, 2002, a partir de estudios mediante trampas parece demostrar la existencia de dos generaciones III-IV, VII-IX), en altitudes de
20 hasta 1200 m.
Que sepamos, los estadios preimaginales son desconocidos.
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Cuenca, Villalba de la Sierra, 40°14’12.42’’N 2°05’23.30’’O, 1000 m, 27.VI.2009, 1♀ sobre Juniperus sabina. Guadalajara, Terzaga, 40°42′47”N, 1°55′01”O, 1347 m, 23.VII.2008, 3♂♂, 2♀♀ sobre J. sabina. Huelva, Matalascañas, 20.VI.2012, 2♂♂, 1♀ sobre J. phoenicea. Teruel, Bezas, 40°19′88′′N, 1°19′74′′O, 1163 m, 23.VII.2008, 1♂ sobre J. sabina, San Blas, 40°21′63”N, 1°10′03”O, 910 m, 22.VII.2008, 6♂♂, 3♀♀ sobre J. sabina.
Coniopteryx (Coniopteryx) borealis Tjeder, 1930 (Figs. 176-178, 302, 303)
Coniopteryx borealis Tjeder, 1930
Tjeder, 1930: 206 (d); Monserrat, 1977c: 139 (list), 142 (list), 1978a: 66 (bio, dis), 1979a: 415 (bio, dis); Monserrat & Marín, 1992: 283 (bio); Marín, 1994: 260-297 (bio, dis, fen); Monserrat & Triviño, 2013: 20 (list); Letardi et al., 2013: 36 (list).
Coniopteryx (Coniopteryx) borealis Tjeder, 1930
Monserrat, 1977c: 139 (rf), 142 (dis, list, mf), 1978b: 183 (bio, dis); Aspöck et al., 1980: 148 (bio, mf, tx, rf); Monserrat, 1984a: 147 (dis, tx), 1984b: 106 (bio, dis), 1984c: 161 (bio, dis), 1984d: 42 (bio, dis), 1984e: 175 (bio, dis), 1984g: 29 (bio,
dis), 1985a: 95 (bio, dis), 1985b: 89 (bio, dis), 1985d: 131 (bio, dis); Díaz-Aranda et al., 1986a: 1125 (bio, dis), 1986b: 1142 (bio, dis); Monserrat, 1987a: 142 (bio, dis); Monserrat & Díaz-Aranda, 1987: 183 (bio, dis), 1988a: 93 (bio, dis); Díaz-Aranda & Monserrat, 1988b: 226 (bio, dis); Monserrat & Díaz-Aranda, 1989: 54 (bio, dis); Marín & Monserrat, 1989: 296 (bio, dis), 1990: 227 (bio, dis); Monserrat & Díaz-Aranda, 1990: 43 (bio, dis); Marín & Monserrat, 1991b: 183 (bio, dis); Monserrat et al., 1994: 58 (bio, dis); Marín & Monserrat, 1995a: 44 (bio, dis), 1995b: 119 (bio, dis); Monserrat, 1995: 120 (bio, dis), 1996a: 15 (list); H. Aspöck et al., 2001: 184 (rf); Monserrat, 2002: 11 (bio, dis), 2005a: 74 (bio, dis), 2013: 300 (bio, dis); Canbulat, 2013: 243 (rf); Monserrat et al., 2013: 54 (bio, dis); Monserrat & Triviño, 2013: 20 (list), 118 (dis).
Especie ampliamente conocida de Europa, Turquía, Cáucaso, Marruecos y Túnez, hasta Azerbaihán, Georgia e Irán. Muy eurioica,
y generalmente citada sobre planifolios, normalmente caducifolios (Quercus, Crataegus, Carpinus, Prunus, Citrus, Corylus, Populus, Betula, Alnus, Castanea, Ulmus, Ceratonia, Pistacea, Acacia,
Fraxinus, Syringa, Salix, Olea, Lycium), a veces sobre frutales, vegetación de rivera, y muy escasamente sobre coníferas (Pinus, Cupressus, Juniperus), entre IV-VIII, en altitudes de hasta 1250 m. En nuestra fauna ha sido ampliamente citada, incluyendo Beleares (Menorca),
recolectada sobre robles, avellanos, alisos, abedules, hayas, quejigos, frutales, chopos, algarrobos, coscojas, encinas, pistáceas,
vegetación de rivera, olmos, acacias, fresnos, olivos, etc., entre IV-XII, Monserrat (2002) demuestra que esta especie puede mantener actividad durante el invierno en fase de imago en zonas costeras y/o meridionales,
y ha sido recolectada en altitudes que oscilan entre 10-1320 m.
Que sepamos, los estadios preimaginales son desconocidos.
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Baleares, Menorca, Shangri la, 25. IX. 1997, 1♀, D. Ventura (DV). Barcelona, Cabrils, 3.VII.1998, 1♂ a la luz, M. Carles-Tolrá, 4.VII.1998, 1♀, M. Carles-Tolrá. Cádiz, Barbate, Caños de Meca, 22.V.2004, 1♂ sobre Olea europaea, 20.VI.2012, 2♂♂ sobre Pistacea lentiscus, Chiclana, Pinar del Hierro, 20.VI.2012, 1♂ sobre P. lentiscus, Estación de La Almoraima, 25.V.2012, 4♂♂, 3♀♀ sobre Quercus canariensis, 21.VI.2012, 5♂♂, 6♀♀ sobre Q. canariensis, Las Cañillas, 21.VI.2012, 1♂ sobre Q. suber, Pinsapar de Grazalema, 3.V.2005, 1♂ sobre Q. rotundifolia. Granada, La Herradura, Cerro Gordo, 27.VII.2013, 2♂♂ sobre P. lentiscus, 17.VIII.2013, 5♂♂, 5♀♀ sobre P. lentiscus, 28.VII.2016, 1♂ a la luz.. Málaga, Las Asomadillas, 25.V.2012, 2♂♂, 1♀ sobre P. lentiscus, 1♂ sobre Q. rotundifolia. Tarragona, La Selva del Camp, 41°13′07.67′′N, 1°08′36.94′′E, 224 m, 21.VIII.2008, 1♂, 1♀, 15.IX.2008, 1♂, 1♀, todos sobre Citrus clementina, N.Cañellas, Tortosa, 10 m, 24.VI.2003, 6 ej., A. Gruppe (AG). Zaragoza, Caspe, 110 m, 24.VI.2003, 1 ej., A. Gruppe (AG).
Coniopteryx (Coniopteryx) parthenia (Navás & Marcet, 1910) sensu Meinander, 1972a (Figs. 167-169, 292-294).
Deasia parthenia Navás & Marcet, 1910 sensu Meinander, 1972a
Navás & Marcet, 1910: 151 (d); Navás, 1910a: 46 (dis, rf), 1924a: 245 (dis, mf); Lacroix, 1924: 55 (rf); Navás, 1925a: 123 (dis, mf); Enderlein, 1930: 103 (rf), 110 (rf); Monserrat, 1985c: 240 (list); Günther, 1993: 168 (rf, tx); Monserrat & Triviño, 2013: 20 (list); Aspöck & Aspöck, 2014: 8 (rf, tx).
Coniopteryx (Coniopteryx) parthenia (Navás & Marcet, 1910) sensu Meinander, 1972a
Meinander, 1972a: 245 (bio, dis, mf); Monserrat, 1977a: 167 (bio, dis), 1978b: 183 (bio, dis); Aspöck et al., 1980: 149 (bio, mf, tx, rf); Meinander, 1981: 107 (dis); Monserrat, 1984a: 147 (dis, tx), 1984b: 106 (bio, dis), 1984c: 161 (bio, dis), 1984d: 42 (bio, dis), 1984e: 175 (bio, dis), 1984g: 29 (bio,
dis), 1985a: 95 (bio, dis), 1985b: 89 (bio, dis), 1985d: 131 (bio, dis); Díaz-Aranda et al., 1986a: 1125 (bio, dis), 1986b: 1142 (bio, dis); Monserrat, 1987a: 142 (bio, dis); Monserrat & Díaz-Aranda, 1987: 182 (bio, dis), 1988a: 93 (bio, dis); Díaz-Aranda & Monserrat, 1988a: 120 (bio, dis), 1988b: 226 (bio, dis), 1988c: 224 (bio, dis); Monserrat & Díaz-Aranda, 1989: 54 (bio, dis); Marín & Monserrat, 1989: 296 (bio, dis), 1990: 227 (bio,dis); Monserrat & Díaz-Aranda, 1990: 43 (bio, dis); Marín & Monserrat, 1991b: 182 (bio, dis); Monserrat et al., 1994: 57 (bio, dis); Marín & Monserrat, 1995a: 44 (bio, dis), 1995b: 118 (bio, dis); Monserrat, 1995: 120 (bio, dis), 1996a: 15 (list), 2002: 11 (bio, dis), 2005a: 73 (bio, dis), 2013: 300 (bio, dis); Monserrat et al., 2013: 54 (bio, dis); Monserrat & Triviño, 2013: 20 (list), 119 (dis).
Coniopteryx parthenia (Navás & Marcet, 1910) sensu Meinander, 1972a
Monserrat, 1977a: 22 (list), 1977c: 142 (list), 1978a: 66 (bio, dis), 1979a: 415 (bio, dis); Monserrat & Marín, 1992: 282 (bio); Marín, 1994: 260-297 (bio, dis, fen).
Por la falta de definición en sus estructuras genitales, desde su descripción (Navás & Marcet, 1910: 151), la posición taxonómica de esta especie ha sido bastante compleja y problemática, no sólo en cuanto a la validez del
género en el que fue descrito (Deasia), basado en aberrantes caracteres de venación alar (sinonimizado a Coniopteryx por Meinander, 1972a) y de la propia especie (redefinida 62 años después por Meinander, 1972a), sino por su relación con alguna de las otras especies europeas de este género (en particular Coniopteryx pygmaea Enderlein, 1906), de las que a partir de multitud de datos bibliográficos (especialmente algunos dados en el centro y norte de Europa), bien
de naturaleza faunística, morfológica o biológica, parecían estar refiriéndose al menos a dos especies distintas: una claramente
asociada a planifolios, otra sobre coníferas (ver Meinander, 1972a), de hecho autores como Tjeder (1931), Killington (1936), Zelený (1961a), Meinander (1962a), Kis (1964a) o Kis et al. (1970) dibujaban como genitalia de C. pygmaea la que resultó ser genitalia de C. parthenia. Fue Meinander (1972a) quien la definió y revalidó, y quien puso fin a esta mezcla de datos existentes entre esta especie y otras, y en particular
C. pygmaea Enderlein, 1906, una de las especies de esta familia más citada en la bibliografía y especie a la que asignó mucha de la información y citas
dadas anteriormente sobre planifolios, restringiendo su distribución geográfica al centro de Europa, y a C. parthenia las dadas sobre coníferas, con una amplia distribución en Europa, norte de África, Oriente Medio y Asia occidental, describiendo
la morfología y la genitalia de ambas especies. Desde entonces fueron consideradas especies distintas y válidas, y fueron
poco a poco corrigiéndose los errores (Greve, 1974; Aspöck et al., 1980; Plant, 1991; Liu & Yang, 1998, etc.), habiendo sido citadas en infinidad de artículos faunísticos.
Como ocurrió en la fauna europea en general, la misma confusión se dio entre las citas antiguas de nuestra fauna (ver comentarios
más adelante de ambas especies y apartado final de citas dudosas o erróneas) agravada, en este caso, por la falta de adecuados
criterios taxonómicos anotados anteriormente en la obra de L. Navás quien, como al inicio hemos referido, no usaba la genitalia
para la identificación de sus ejemplares y citó profusamente en nuestra fauna varias especies de este género (Coniopteryx), mezclando especies sin orden ni concierto (Monserrat, 1984a; Monserrat et al., 1994; Monserrat, 2010) y generando consecuentemente posteriores referencias de varias especie en nuestra fauna (ver apartado anterior sobre la
problemática de las citas antiguas y el apartado final de especies dudosamente citadas), y según hemos anotado, por la enorme
cantidad de errores de identificación detectados en sus citas, Monserrat (1984a) propuso que las citas de esta familia dadas por este autor, al menos en nuestra fauna, no debían ser consideradas.
Aunque es probable que muchas de sus citas de la especie a la que ahora nos referimos pudieran haber sido “acertadas” o pudieran
presentar la especie que ahora tratamos (especialmente las de ejemplares recolectados en coníferas), es muy probable que alguna
de las especies citadas en nuestra fauna como Coniopteryx tineiformis y/o Coniopteryx pygmaea, podrían pertenecer a la especie que tratamos (las citas dadas sobre ejemplares recolectados en coníferas, como la cita de
Coniopteryx tineiformis de Navás, 1914c: 41: “abundante en los pinos” o de Navás, 1917a: 42: “frecuente en los árboles, en especial Pinus uncinata” son significativas al respecto), pero esto solo puede sugerirse, a no ser de haberse conservado y estudiado los ejemplares
(ver más adelante comentarios en C. tineiformis y de C. pygmaea en el apartado final de especies errónea o dudosamente citadas). Sin embargo, la realidad es otra, pues Monserrat (1984a) estudió los ejemplares que se conservaron en la colección L. Navás, y entre ellos (salvo el holotipo), los que fueron identificados
por Monserrat (1984a) como pertenecientes a C. parthenia, ninguno habían sido identificado y/o citado por Navás con este nombre, sino como pertenecientes a otras especies como C. tineiformis, C. pygmaea, e incluso como Conwentzia psociformis. Por todo ello es mejor no considerar estas citas antiguas.
Volviendo a la identidad de Coniopteryx pygmaea y a su relación con la especie que tratamos, sus caracteres de morfología y especialmente su genitalia masculina estaban
perfectamente definidos desde Enderlein (1906: fig. 55; 1908: fig. 9) y, como hemos anotado, ha sido citada y reproducida en numerosas ocasiones (ver Meinander, 1972a, 1980 o Aspöck et al., 1980). Pues bien, con aparentemente idénticas características morfológicas se describieron posteriormente dos especies
europeas: Coniopteryx transsylvanica Kis, 1964 de Rumanía y Coniopteryx hoelzeli Aspöck, 1964 de Austria, especies a las que ahora nos referiremos. Por la importancia y consecuencias posteriores de esta
decisión, conviene detenerse y manifestar nuestra opinión y argumentos.
Los ejemplares de la Serie Tipo de Coniopteryx pygmaea (Enderlein, 1906) estaban constituidos por tres Sintipos: dos machos (Alemania: Berlín, Rahnsdorf, 15.V.1890, Grünau, 10.VIII.1902) y una
hembra (Alemania: Berlín, Straussberg, 4.VIII.1901). Aunque a diferencia de otras colecciones entomológicas alemanas, las
del Museums für Naturkunde der Humboldt-Universität zu Berlin milagrosamente no sufrieron demasiados daños durante los terribles
destrozos originados por los conflictos bélicos en Europa, especialmente durante las dos Guerras Mundiales (Weidner, 1972), y en particular nos referimos a los Ejemplares Tipo de neurópteros (Horn et al., 1990; Ohl, 2002). Aún así Meinander (1972a) no pudo localizar estos ejemplares, pero que en base a la descripción y los dibujos originales de Enderlein (1906: 201, figs. 6, 36, 55, 58, 61; 1908: 9, fig. 9) asignó a esta especie lo que se había descrito como Coniopteryx transsylvanica Kis, 1964, especie descrita de Moneasa en Rumanía (Kis, 1964b) y propuesta como sinonimia de la siguiente especie por el propio Kis (1965c), y como Coniopteryx hoelzeli Aspöck, 1964 (descrita en base a un ejemplar recolectado en un bosque mixto de robles y hayas en Carintia, en el sur de Austria),
asumiendo o proponiendo las consecuentes sinonimias. En base a ello, lo anteriormente citado (sobre coníferas) o descrito
como segunda especie mezclada en la bibliografía con ella lo asignó a Coniopteryx parthenia, tanto por la abundancia de citas morfológicas, de genitalia y de biología, como por el estudio de su ejemplar tipo (♀),
que en buen estado se conserva en el Museo de Zoología de Barcelona (Meinander, 1972a), y cuya genitalia es muy característica (Figs. 292-294). Como hemos indicado, desde entonces ambas especies han sido abundantemente citadas en la bibliografía (conocemos más de
140 artículos y trabajos que la citan, de los que 43 afectan a nuestra fauna).
Durante tres décadas todo parecía estar aclarado desde el punto de vista taxonómico, hasta que Kleinsteuber (1974) y Günther (1991) sugieren que ambas especies (Coniopteryx pygmaea y Coniopteryx parthenia) pudieran ser sinónimas, y más tarde Günther (1993) descubre los sintipos de Coniopteryx pygmaea, los estudia, y redescribe su genitalia masculina en base a nuevos elementos (no reflejados en la descripción original).
Estos tres ejemplares (sintipos), 2 ♂♂, 1♀, están montados en preparaciones microscópicas y uno de los machos carece de abdomen,
designando al otro ejemplar macho como lectotipo (Berlin, Rahnsdorf, 15.5.1890 Tetens S.).
En base a estos estudios, Günther (1993) concluye que el lectotipo elegido poseía caracteres que lo asocian a lo citado en Coniopteryx parthenia (Navás & Marcet, 1910) (sensu Meinander, 1972a), proponiendo a esta especie como sinónima posterior de Coniopteryx pygmaea Enderlein, 1906, liberando pues a Coniopteryx hoelzeli de ser considerada, como hasta entonces, de ser sinónima posterior de Coniopteryx pygmaea (sensu Meinander, 1972a). En base a nuestro criterio y opinión personal, y en base a elementos exclusivamente científicos y objetivos, consideramos
oportuno comentar y discutir esta propuesta, y es el momento de exponerlos.
Sinceramente nos parece extraordinariamente extraño que Enderlein (1906), que tanta delicadeza y precisión demostró en la elaboración de sus descripciones y sus dibujos (oponión también compartida
y alabada por Tjeder, 1931; Killington, 1936; Meinander, 1972a; Günther, 1993; Aspöck et al., 1980; Aspöck & Aspöck, 2003, etc.), no hubiera reparado, detectado y descrito unas estructuras tan esclerificadas y conspicuas como las que hallamos
en la genitalia masculina de Coniopteryx parthenia (sensu Meinander, 1972a) (Figs. 167, 168) al describir Coniopteryx pygmaea Enderlein (1906: 201, fig. 55; 1908: fig. 9), y que Günther (1993: figs. 1,2) describe del lectotipo, y sobre todo que su genitalia masculina se encuentre tan destrozada como Günther (1993: fig. 1) anota, cuando las figuras de Enderlein (1906: fig. 55; 1908: fig. 9), no lo reflejan en absoluto, y nos sorprende que los elementos anotados entre uno y otro autor sean
tan dispares (Enderlein, 1906: 201, figs. 6, 36, 55, 58, 61; 1908: 9, fig. 9; Günther, 1993, figs. 1, 2).
En lo que respecta a la genitalia masculina, Enderlein (1906: 201) describe con toda claridad los parámeros de C. pygmaea: “die beiden Penishälften sonst ohne Anhänge oder Äste”, y sinceramente tampoco se nos asemeja mucho a lo mencionado por Günther (1993). Por ello, no estamos en absoluto de acuerdo con Günther (1993) cuando refiriéndose a la descripción de Enderlein (1906) anota: “La ilustración “incompleta” de la genitalia del ♂ de C. pygmaea por parte del autor, donde no quedan reflejadas las regiones más apicales de los parámeros (menos esclerotizados) es con
seguridad el causante de la decisión errónea de Meinander”. Nosotros nunca hemos observado entre los cientos de ejemplares
machos estudiados de Coniopteryx parthenia (sensu Meinander, 1972a), que tales zonas más apicales de los parámeros estén menos esclerotizadas (o menos pigmentadas) (Figs. 167, 168), ni que la más ventral sea recta y/o levemente acodada al resto, como ocurre en C. pygmaea sensu Enderlein, 1906 (Figs. 170, 171), sino continua y uniformemente recurvada (Figs. 167, 168), y que la más dorsal sea membranosa, y no digamos espatulada, elementos que expone para ambas estructuras Günther (1993: figs. 1, 2), y jamás así lo hemos visto representada en la información publicada sobre lo que era o resultó ser la genitalia
masculina de esta especie (Tjeder, 1931; Killington, 1936; Zelený, 1961a, Meinander, 1962a; Kis, 1964a; Kis et al., 1970; Meinander, 1972a; Liu & Yang, 1998; Sziráki, 2011). Tampoco vemos en el ejemplar (lectotipo) estudiado y reconstruido por Günther (1993: figs. 1, 2) ninguna línea de fractura que vincule los parámeros y sus procesos apicales, completamente diferentes a lo
conocido en otras especies europeas del subgénero, inclinándonos a sugerir si no es un artefacto en la preparación microscópica
estudiada.
Por otra parte, reconstruyendo los fragmentos del hipandrio del ejemplar (lectotipo) estudiado y haciendo coincidir las líneas
de fractura anotadas por Günther (1993: fig. 1), la incisión media resultante es mucho más en “U cerrada”, como ocurre en C. pygmaea sensu Enderlein, 1906 (Fig. 172) que en “V abierta”, como ocurre en C. parthenia (sensu Meinander, 1972a) (Fig. 169), hecho que no coincide con lo reconstruido por Günther (1993: fig. 2). En cualquier caso, y como también ocurre en Coniopteryx pygmaea (sensu Meinander, 1972a), el margen caudal del hipandrio es muy variable en sus procesos, siendo los más centrales y su incisión media desde redondeados
(Aspöck et al., 1980: 56, fig. 285; Plant, 1991: 114, fig. 22) a marcadamente angulosos (Meinander, 1962a: 23, fig. 20; 1972a: 246, fig. 175E; Liu & Yang, 1998: 188, fig. 2C, etc.).
Tampoco entendemos que, existiendo un ejemplar ♀ (paralectotipo) montado en preparación microscópica (Alemania: Berlín, Straussberg,
4.VIII.1901), no hubiera sido estudiado y descrito por Günther (1993) para argumentar y apoyar su propuesta, máxime cuando la genitalia femenina de C. parthenia (sensu Meinander, 1972a) ya ha había sido descrita en detalle (Meinander, 1972b; Sziráki, 1992a, 1992b, 1992c) y cuya genitalia es muy característica (Figs. 292-294).
Al margen de la morfología genital, nos parece muy curioso que Meinander (1972a: 249) no pudiera localizar estos ejemplares al realizar su revisión mundial (“ENDERLEIN′s types have unfortunately not been
traced”), y que el citado museo no tuviera registro de su préstamo a P. Ohm, según alegan Günther (1991: 162, 1993: 168) “fueron prestados del museo al Dr. Peter Ohm, Kiel, desde el 22.1.1968”, o Aspöck & Aspöck (2014: 8) “Estos habían sido prestados por el Museo de Berlín de Peter Ohm (1922-2001) y mantenidos por él durante muchos años”,
y no deja de ser sorprendente que este hecho no hubiera sido comentado entre ambos autores (Ohm y Meinander) que estaban en
permanente contacto y así lo refleja Meinander (1972a) a lo largo de todo el texto de su obra en el material estudiado o citado, y expresamente lo menciona (Meinander, 1972a: 7) en los agradecimientos a instituciones y colegas.
Además, la alegación al comentario Tjeder (1931) cuando supuestamente describía la genitalia de Coniopteryx pygmaea: “The specimens which I refer to this species do not agree very well with ENDERLEIN′s description of the aedeagus...”, posteriormente
comentado por Meinander (1972a), no puede aplicarse como argumento, ya que entonces Tjeder (1931) estaba comparando ejemplares que creía de la misma especie, pero que eran dos especies diferentes: Coniopteryx pygmaea (sensu Enderlein, 1906) y Coniopteryx parthenia (sensu Meinander, 1972a), por entonces confundidas. También Günther (1993) alega que Meinander (1972a) no cita el locus typus de Coniopteryx parthenia, cosa que sí hace (Meinander, 1972a: 248: Distribution: Germany Berlin).
Nos resulta sorprendente y verdaderamente audaz que K.K. Günther, que sepamos, con escasa experiencia previa (salvo un artículo
faunístico, Günther, 1991) o posterior en el campo de la familia que tratamos (Oswald, 2013b) abordara un trabajo taxonómico de este calado, y sinceramente preferimos seguir y confiar en la experiencia y el criterio
de G. Enderlein y M. Meinander. Más nos sorprende aún el no haber recibido, hasta la fecha de hoy, contestación del Museo
de Berlín sobre la información solicitada de estos lectotipos para obtener personalmente otras averiguaciones.
Por último, las dudas sobre lo acertado de esta designación se acrecientan al tener en cuenta los datos conocidos sobre la
biología de ambas especies: durante más de tres décadas Coniopteryx parthenia (sensu Meinander, 1972a) ha sido una especie abrumadoramente asociada a coníferas [aunque no se especifica claramente en la descripción original
(Navás & Marcet, 1910), la asociación de esta especie con coníferas se aprecia e intuye desde ella, dada la vegetación de la zona (Marcet, 1948-1953; Panareda Clopés & Nuet Badia, 1993-1994; Panareda Clopés, 2011), donde entre bosques mediterráneos, y en función del tipo de suelo, se desarrollan bosques/bosquetes de Pinus nigra, P. pinea y P. halepensis y que, a pesar de las heladas de enero de 1985 que afectaron en particular a las especies mediterráneas termófilas, como
Pinus halepensis y de los incendios de agosto de 1986, aún perduran en la actualidad], y practicamente todos los datos bibliográficos existentes
la relacionan con coníferas, a las que acompaña en su distribución y repoblación, especialmente Pinus, Abies, Juniperus y Cedrus, también ha sido muy puntualmente citada sobre otras plantas (Tamarix, Ilex, Ficus, Erica, Fagus, Tetraclinis o Quercus), también en nuestra fauna [como datos ilustrativos mencionemos que Monserrat & Marín, 1992 recogen la información existente hasta la fecha sobre las plantas donde se habían recolectado los ejemplares ibéricos de
esta familia, y de los 1758 ejemplares estudiados de C. parthenia (sensu Meinander, 1972a) recolectados sobre diversas plantas, solo 27 no habían sido recolectados en coníferas, de los 2265 estudiados por nosotros
hasta ahora de C. parthenia (sensu Meinander, 1972a) recolectados sobre diversas plantas, solo 47 no habían sido recolectados en coníferas, y de los 302 nuevos ejemplares ahora
citados sobre diferentes plantas, solo 1 no ha sido recolectado sobre coníferas]. Por el contrario, nada de esa vinculación
con coníferas se menciona en la descipción de Coniopteryx pygmaea (ver Enderlein, 1906) (sensu Meinander, 1972a), que ha sido mayoritariamente citada sobre planifolios (Quercus, Salix, Populus, Crataegus, Prunus, Corylus, Malus, Fagus, Acer, Alnus, Carpinus o Tilia) (datos que coinciden con los del tipo de Coniopteryx hoelzeli).
Consideramos que sería relativamente sencillo que, de aceptar esta sinonimia, se provocara el transvase de información de
todas las citas existentes sobre coníferas de C. parthenia (sensu Meinander, 1972a) a C. pygmaea (sensu Günther, 1993), especie (C. parthenia) de la que Aspöck & Aspöck (2014: 8) concluyen: “is the species confined to coniferous trees and occurs very abundantly in all parts of Europe, even in the
north of Scandinavia, moreover in Morocco, Asia minor, Northern, Central, and Eastern Asia” pero, de seguir este criterio,
y conociendo que C. pygmaea es una de las especies más citada en la bibliografía (sólo superada en citas por C. tineiformis), sería extremadamente arriesgado transvasar la ingente cantidad de citas sobre planifolios de esta especie (C. pygmaea) a C. hoelzeli (sensu Günther, 1993), cuando Aspöck & Aspöck (2014: 8), tras defender el criterio de Günther (1993), concluyen sobre C. hoelzeli: “it is rare, usually found in single specimens and known only from a few spots in the south of Central Europe, in Eastern
Europe and in the Far East... Most probably it does not occur in the north of Central Europe”. De ser así, ¿a qué otra especie
habría que asignar todas estas de citas?
Igual problemática se genera con la morfología genital femenina y/o los estadios preimaginales descritos para estas especies
en base a material recolectado sobre uno y otro tipo de plantas. ¿A qué especie habría que asignar estas de descripciones?
Por último, y sobre este particular, estando perfectamente definidas ambas especies desde Meinander (1972a), son cientos las citas asignadas a una u otra especie desde entonces. En lo que respecta a C. pygmaea (sensu Meinander, 1972a) conocemos de la bibliografía general más de 180 artículos y trabajos que la citan [a los que habría que sumar 21 artículos
y trabajos que citan a C. hoelzeli (sensu Meinander, 1972a) y 5 artículos y trabajos que citan a C. transsylvanica (sensu Meinander, 1972a)], y con respecto a C. parthenia (sensu Meinander, 1972a) conocemos más de 140 artículos y trabajos que la citan, de los que 43 afectan a nuestra fauna, por lo que, en todo caso
esta especie (C. parthenia sensu Meinander, 1972a), podría considerarse Nomina conservanda por la Comisión Internacional de Nomenclatura Zoológica. En definitiva, salvo recuperar una especie (C. hoelzeli) de la lista de sinonimias en la que había sido asumida en las últimas décadas, por todo lo demás, esta propuesta de Günther (1993), solo traería confusión.
Tenemos suficientes argumentos pues como para dudar y no aceptar la propuesta de Günther (1993), y ante todos estos elementos, no asumimos ni podemos aceptar las propuestas de Günther (1993), Aspöck et al. (2001) o Aspöck & Aspöck (2014) que ahora hemos rebatido y discutido. Por todo ello, seguimos manteniendo el criterio de Enderlein (1906, 1908) y Meinander (1972a), a pesar de las veladas críticas vertidas a nuestras opiniones sobre esta decisión (Aspöck & Aspöck, 2014: 8 “The time has come to stop the absolutely unjustified and unnecessary confusion”). Sin duda compartimos los afectos a
nuestro admirado Herbert Hölzel [Aspöck & Aspöck, 2014: 9 “We cannot accept that the species which was named in honour of Herbert Hölzel (1925-2008) is being pushed into a clearly
wrong synonymy!”], pero no son los criterios y testimonios emocionales los ahora válidos, sino los objetivos y científicos,
y como hemos indicado, no nos convencen y creemos han sido ahora suficientemente argumentados. En la Ciencia no hay ganadores
ni perdedores, sólo opiniones para enriquecerla-enriquecernos. Ya decía Epicuro. “En una disputa filosófica (científica), gana más el que más pierde porque es el que más aprende”.
Por todo ello, ex toto, mantenemos nuestro criterio sobre la validez de la/s especie/s que tratamos, tanto de Coniopteryx parthenia (sensu Meinander, 1972a) como de Coniopteryx pygmaea (sensu Enderlein, 1906 y Meinander, 1972a), y ahora manifestamos que sí era necesaria y justificada declarar nuestra opinión y decisión sobre estas especies, ajustándonos
a lo que sobre ellas hemos argumentado, citado y conocemos. Retomamos pues Coniopteryx parthenia (sensu Meinander, 1972a).
Elemento paleártico occidental, ampliamente conocido desde el norte de África a Mongolia. Está fuertemente asociado y mayoritariamente
citado sobre coníferas, a las que acompaña en su distribución, especialmente Pinus y Cedrus, también puntualmente citado sobre otras plantas (Tamarix gallica, Tetraclinis articulata o Quercus ilex), entre IV-XI, en altitudes de 100 hasta 2020 m. En nuestra fauna ha sido ampliamente citada, incluyendo Baleares (Formentera),
mayoritariamente sobre coníferas (Pinus, Abies, Juniperus, Cedrus), excepcionalmente sobre Tamarix, Ilex, Ficus, Erica, Fagus, Tetraclinis y Quercus, entre III-XI, en altitudes que oscilan entre 10-2100 m. Con frecuencia es extraordinariamente abundante.
Como hemos citado con la confusión de especies entre los imagos, los estadios preimaginales de especies descritos por Curtis (1834, pl. 528), Vine (1895: 265), Withycombe (1923: 582), Lacroix (1924: 74), Killington (1936: 203), Genay (1953: 23), Rousset (1956a: 1, 1956b: 1, 1958: 1), Ghilarov (1962: 412), Rousset (1966: 8, 1969: 108) o Meinander (1972a: 31), aunque han sido mayoritariamente descritos como pertenecientes a C. tineiformis o C. pygmaea, es probable que mayoritariamente puedan pertenecer a la especie que tratamos (Meinander, 1972a; Greve, 1974).
La genitalia de un único ejemplar (♀) recolectado en Cerdeña permite asignarlo a esta especie, según lo anotado por Meinander (1972a) y Sziráki (1992a), resultando una especie nueva para esta isla.
NUEVO MATERIAL Y MATERIAL COMPLEMENTARIO ESTUDIADO: ESPAÑA: Álava, Zuia, Gorbeia P.N., 30TWN1362, 800 m, 23.VI.2007, 1♀ sobre Pinus nigra, S. Pagola. Asturias, Luarca, 2.VII.2011, 1♀ sobre P. sylvestris, V. Triviño. Barcelona, Cabrils, 1.VII.1998, 1♀ a la luz, M. Carles-Tolrá. Cádiz, Barbate, Caños de Meca, 20.VI.2012, 1♂ sobre P. pinea, Pinsapar de Grazalema, 3.V.2005 1♂ sobre Abies pinsapo, 26.V.2012, 3♂♂, 4♀♀ sobre A. pinsapo, 22.VI.2012, 1♂, 3♀♀ sobre A. pinsapo, Playa de los Lances, 21.VI.2012, 1♂ sobre P. pinea. Cuenca, Huélamo, Puerto del Cubillo, 1617 m, 22.VII.2008, 1♀ sobre P. sylvestris. Granada, Acebuche, 287 m, 13.IV.2014, 2♂♂, 3♀♀ sobre P. halepensis, Aldeire, 30SVG9410, 1661 m, 8.V.2011, 1♀ sobre P. pinaster, Calahonda, 30SVF66, 20 m, 8.VII.2013, 5♂♂, 5♀♀ sobre P. halepensis, Collado de la Sabina, 30SVG6207, 2050 m, 30.VI.2011, 58♂♂, 29♀♀sobre P. sylvestris, 25.VIII.2011, 1♂ sobre P. sylvestris, 10.VII.2012, 2♂♂, 5♀♀ sobre P. sylvestris, 10.VII.2013, 5♂♂, 14♀♀ sobre P. sylvestris, 27.VII.2013, 1♂, 3♀♀ sobre P. sylvestris, Ferreira, 30SVG9513, 1356 m, 8.V.2011, 1♂, 1♀ sobre P. halepensis, Gualchos, 30SVF6462, 30 m, 28.VI.2011, 7♂♂ sobre P. halepensis, Hoya de la Mora, 30SVG6506, 2500 m, 30.VI.2011, 5♂♂, 6♀♀ sobre P. sylvestris, 25.VII.2013, 1♂ sobre P. halepensis, La Herradura, Cantarriján, 30SVF36, 10 m, 12.VI.2012, 2♂♂, 3♀♀ sobre P. halepensis, F. Acevedo, 9.VII.2013, 1♀ sobre P. halepensis, La Herradura, Cerro Gordo, 6.VIII.2011, 2♂♂ a la luz, 27.VIII.2011, 1♂ a la luz, 17.VIII.2013, 4♀♀ sobre P. halepensis, La Herradura, Punta de la Mona, 30SVF36, 200 m, 7.IV.2007, 2♂♂, 1♀ a la luz, 12.VIII.2010, 1♂ a la luz, Lugros, 19.VII.2011,
1♂ sobre P. halepensis, Presa de Rules, 280 m, 13.IV.2014, 2♂♂, 3♀♀ sobre P. halepensis, Valle Río Izbor, 36°52′52”N, 3°27′11”O, 300 m, 17.VI.2010, 1♂, 1♀ sobre P. halepensis. Huelva, Arroyo Julianejo, 24.V.2012, 7♂♂, 4♀♀ sobre P. pinea, Matalascañas, 20.VI.2012, 11♂♂, 2♀♀ sobre P. pinea, Punta Umbría, 24.V.2012, 1♂ sobre P. pinea, 20.VI.2012, 9♀♀ sobre P. pinea. Islas Baleares, Menorca, Cala Turqueta, 23.VII.2008, 1♀ sobre P. halepensis, F. Acevedo. Madrid, El Escorial, Abantos, Fuente del Cervunal, 1.700 m, 3.VII.2007, 1♂, 3♀♀ sobre P. sylvestris, 24.VII.2007, 2♀♀ sobre P. sylvestris, 7.VI.2008, 3♂♂, 1♀ sobre P. sylvestris, 21.VI.2008, 7♂♂, 10♀♀ sobre P. sylvestris, 2.VII.2008, 3♀♀ sobre P. sylvestris, 31.VII.2008, 1♀ sobre P. sylvestris, 4.VII.2012, 3♀♀ sobre P. sylvestris, 29.VI.2013, 1♂, 2♀♀ sobre P. halepensis, 19.VI.2014, 4♀♀ sobre P. sylvestris, F. Acevedo, Arboreto Luis Ceballos, 29.VIII.2007, 1♀ sobre P. pinaster, 7-21.VI.2008, 1♂ en Trampa de Malaise, 17-31.VII.2008, 1♀ en Trampa de Malaise, 1.VIII-12.IX.2008, 1♂, 1♀ en Trampa de Malaise.
Málaga, Coín, Sa. Alpujata, 3.V.2013, 1♀ sobre P. halepensis, Maro, 2.V.2014, 2♂♂, 3♀♀ sobre P. halepensis, Pinos de Alhaurín, 220 m, 2.V.2014, 2♂♂, 3♀♀ sobre P. halepensis, Torrox, 5.VIII.2010, 2♂♂, 3♀♀ sobre P. halepensis, 25.VIII.2010, 1♂ sobre P. halepensis, Vélez Málaga, 5.VIII.2010, 3♂♂, 1♀ sobre P. halepensis, 25.VIII.2010, 1♂, 1♀ sobre P. halepensis. Murcia, Pinilla, 10.VII.2005, 3♂♂, 2♀♀ sobre P. halepensis. Segovia, Valsaín, 8.VII.2007, 3♀♀ sobre P. sylvestris. Tarragona, La Selva del Camp, 41°13′07.67′′N, 1°08′36.94′′E, 224 m, 20.X.2010, 1♂ sobre Citrus clementina, N. Cañellas. Teruel, Frías de Albarracín, 40°20′43”N, 1°36′78”O, 1465 m, 22.VII.2008, 1♀ sobre Juniperus communis. Zaragoza, Daroca, 17.VI.2007, 1♀ sobre P. halepensis. ITALIA: Sardegna, Olbia Tempio, Telti, Monte Pino, 40°52′15′′N, 9°23′03′′E, 300 m, 20.V.2008, 1♀ sobre P. pinaster.
Coniopteryx (Coniopteryx) tineiformis Curtis, 1834 (Figs. 11-14, 173-175, 301)
Coniopteryx tineiformis Curtis, 1834
Curtis, 1834: 528 (d); Navás, 1908: 129 (dis, mf); Macho Bariego, 1909: 75 (dis); Navás, 1910a: 45, 46, 49, 52, 54 (dis), 1910b: 244 (dis); Beché, 1913: 170 (dis); Navás, 1913a: 85 (dis), 1913b: 76 (dis), 1914a: 35 (dis), 1914b: 215 (dis), 1914c: 41 (bio, dis), 1914d: 190 (dis), 1915: 30, 35, 39, 43, 52, 68, 70, 74, 76 (dis), 1916a: 174 (dis), 1916b: 155 (dis), 1916c: 190 (dis); Bohigas & Sanchez, 1917: 318 (dis); Navás, 1917a: 41 (dis), 1917b: 88 (dis), 1918c: 41 (dis), 1919a: 202 (dis), 1919b: 43 (dis), 1921a: 152 (dis), 1921b: 65 (dis), 1922: 110 (dis), 1923a: 168 (dis), 1923b: 8 (dis), 1924a: 242 (dis, key, mf), 1924b: 26 (dis), 1925a: 121, 122 (dis, key, mf); Stitz, 1927: 18 (rf); Navás, 1928: 102 (dis); 1929a: 33 (dis), 1930: 52 (dis), 1931: 121 (dis); Eglin, 1940: 269 (rf); Vidal y López, 1943: 24 (rf); Berland, 1962: 26 (rf); Monserrat, 1977a: 170 (rf), 1977c: 142 (list); Sziráki, 1979: 181 (rf); Séméria & Berland, 1988: 53 (rf); Monserrat & Marín, 1992: 282 (bio); Ocharan et al., 2012: 246 (rf); Monserrat & Triviño, 2013: 20 (list); Letardi et al., 2013: 36 (list).
Coniopteryx (Coniopteryx) tineiformis Curtis, 1834
Monserrat, 1978b: 183 (bio, dis); Aspöck et al., 1980: 148 (bio, mf, tx, rf); Monserrat, 1984a: 147 (dis, tx), 1984d: 42 (bio, dis), 1984g: 28 (bio, dis), 31 (rf), 1985a: 95 (bio, dis); Marín & Monserrat, 1989: 295 (bio, dis); Monserrat et al., 1994: 57 (bio, dis); Marín & Monserrat, 1995b: 118 (bio, dis); Monserrat, 1995: 120 (bio, dis), 1996a: 15 (list); H. Aspöck et al., 2001: 186 (rf); Monserrat, 2005a: 73 (nt), 2010: 26 (bio, dis); Monserrat et al., 2013: 54 (bio, dis); Monserrat & Triviño, 2013: 20 (list), 119 (dis).
Malacomyza lactea Wesmael, 1836
Wesmael, 1836a: 167 (d).
Coniopteryx lactea (Wesmael, 1836)
Navás, 1905a: 507 (dis), 1905b: 36 (rf); Monserrat & Triviño, 2013: 20 (list).
Coniopteryx tineiformis xaveriana Navás, 1918
Navás, 1918b: 20 (d), 1921b: 65 (dis), 1924a: 243 (dis, mf); Lacroix, 1924: 55 (rf); Navás, 1925a: 122 (dis, mf), 1932: 16 (dis); Monserrat, 1985c: 240 (list); Monserrat & Triviño, 2013: 20 (list).
Coniopteryx tineiformis xaverina Navás, 1918 (lapsus)
Meinander, 1972a: 252 (rf, tx).
Tratamos la especie de esta familia más citada en la bibliografía. La imprecisa descripción de esta especie (Curtis, 1834) estuvo basada en una mezcla de ejemplares pertenecientes a varias especies y géneros distintos. Fue Meinander (1972a) quien entre ellos eligió un ejemplar ♂ al que designó como lectotipo y desde entonces así quedó definida y ha sido mayoritariamente
aceptada.
Como hemos anotado, se trata de una especie profusamente citada en nuestra fauna, aunque como ahora veremos, la mayoría de
estas citas eran incorrectas. Ya hemos indicado que Monserrat (1984a) estudió los ejemplares de la colección L. Navás, identificados y citados como pertenecientes a la especie que tratamos (la
mayor parte de estas citas antiguas, más de 30, sólo de Navás), y en solo dos ejemplares (de Santiago en La Coruña y de el Moncayo en Zaragoza) la identificación era correcta (Monserrat, 1984a; Monserrat et al., 1994; Monserrat, 2010). El resto de estos ejemplares así identificados se correspondían hasta con diez especies distintas (alguno incluso de otros
géneros como Conwentzia o Semidalis), confusiones que eran extensibles a otras especies por él identificadas, por lo que Monserrat (1984a) sugería no considerar las citas de este autor referentes a esta familia. Sobre otras citas antiguas de esta especie que
se han anotado y que no han podido corroborarse, especialmente las dadas en Levante son muy dudosas. Por lo que ahora conocemos,
todas las citas de esta especie fuera de la región ibérica de influéncia eurosiberiana, incluso éstas que no hayan sido comprobadas,
deben ponerse en duda o desestimarse (Monserrat, 1984a, 2005a, 2010).
Especie paleártica occidental (quizás holártica), citada en USA (Alaska, Michigan, Tennessee, Idaho, Virginia) y Canadá (Newfoundland,
Quebec), habitualmente recolectada en medios boscosos eurosiberianos sobre vegetación caducifolia (Corylus, Populus, Fagus, Quercus, Salix, Alnus, Carpinus, Betula, Rubus, Sorbus, Acer, etc.), también sobre vegetación de rivera y en jardines, entre IV-X, en altitudes de 500 hasta 1800 m. En nuestra fauna
tiene una marcada tendencia eurosiberiana, habiéndose confirmado en su tercio septentrional (Lérida, Huesca, Navarra, Zaragoza,
Santander, Asturias, Zamora, Orense y Lugo) (Monserrat, 2005a cuestiona las citas en Mallorca y Monserrat 2010 comenta el historial de las citas en nuestra fauna), y ha sido citada sobre robles, arces, hayas, avellanos, abedules, chopos,
hierba y vegetación de rivera, entre VI-VIII, en altitudes que oscilan entre 60-1687 m. No tenemos ninguna constancia (ni
fiabilidad) de que las citas antiguas de Coniopteryx tineiformis Curtis, 1834 o de Coniopteryx tineiformis xaveriana Navás, 1918 pertenezcan, en realidad, a esta especie (el único ejemplar existente en su colección descrito, identificado
y citado por él en Navás, 1918b como Coniopteryx tineiformis xaveriana, pertenecía, según Monserrat, 1984a, a C. borealis).
Alguno de los estadios preimaginales descritos de esta especie por Curtis (1834: pl. 528, fig. L), Vine (1895: 265), Withycombe (1923: 584), Lacroix (1924: 74) o Killington (1936: 198) pueden estar refiriendose a C. parthenia.
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Asturias, Luarca, 2.VII.2011, 1♂, 1♀ sobre Fagus sylvatica, V. Triviño.
Coniopteryx (Holoconiopteryx) drammonti Rousset, 1964 (Figs. 179-187, 295, 296)
Coniopteryx drammonti Rousset, 1964
Rousset, 1964: 2 (d); Joost, 1973: 152 (rf); Monserrat, 1975: 264 (list); Marín & Monserrat, 1991a: 192 (bio, dis, fen); Monserrat & Marín, 1992: 283 (bio); Sziráki, 1992c: 359 (rf); Marín, 1994: 260-297 (bio, dis, fen); Monserrat, 2010: 28 (rf); Monserrat & Triviño, 2013: 20 (list).
Coniopteryx (Holoconiopteryx) drammonti Rousset, 1964
Monserrat, 1975: 264 (list, mf); Aspöck et al., 1980: 151 (bio, mf, tx, rf); Monserrat, 1984b: 107 (bio, dis), 1984c: 161 (bio, dis), 1984e: 176 (bio, dis); Díaz-Aranda et al., 1986a: 1125 (bio, dis), 1986b: 1143 (bio, dis); Monserrat & Díaz-Aranda, 1987: 183 (bio, dis); Díaz-Aranda & Monserrat, 1988a: 120 (bio, dis); Marín & Monserrat, 1990: 228 (bio, dis), 1991b: 183 (bio, dis); Monserrat et al., 1994: 58 (bio, dis); Marín & Monserrat, 1995b: 119 (bio, dis); Monserrat, 1996a: 15 (list); H. Aspöck et al., 2001: 188 (rf); Monserrat, 2013: 300 (bio, dis); Monserrat & Triviño, 2013: 20 (list), 120 (dis).
Coniopteryx clavata Monserrat, 1975
Monserrat, 1977a: 22 (list), 1977c: 142 (list), 1978a: 66 (bio, dis), 1981c: 182 (nt); Séméria & Berland, 1988: 54 (rf); Monserrat & Triviño, 2013: 20 (list).
Coniopteryx (Holoconiopteryx) clavata Monserrat, 1975
Monserrat, 1975: 259 (d), 1977a: 171 (key, mf), 172 (bio, dis); Aspöck et al., 1980: 152 (tx); Monserrat, 1981c: 182 (nt); Monserrat & Triviño, 2013: 20 (list).
Especie conocida del sur y centro de Europa, Marruecos, Turquía, Georgia e Irán. Generalmente recolectada sobre Quercus, pero también sobre Juniperus, Fagus, Acer, Crataegus, Celtis, Pinus y Carpinus, entre V-IX, en altitudes de hasta 2031 m. En nuestra fauna está ampliamente citada, especialmente en su región de influencia
mediterránea, y ha sido citada sobre encinas, robles, quejigos, hayas, arces, sabinas, alnueces, cornillos y pistáceas, entre
V-IX, en altitudes que oscilan entre 40-2000 m.
Que sepamos, los estadios preimaginales son desconocidos.
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Barcelona, Caldes de Montbui, 10.VII.1998, 1♂ a la luz, C. Torras, 1.X.1998, 1♂ a la luz, C. Torras. Cádiz, Hozgarganta, Los Alcornocales, 21.VI.2012, 1♂, 1♀ sobre Quercus canariensis, La Marchenilla, 25.V.2012, 1♂ sobre Olea europea. Granada, Cáñar, 18.VII.2011, 1♂, 1♀ sobre Q. pyrenaica, El Dornajo, 37°07’52.22’’N, 3°26’57.43’’O, 1750 m, 11.VII.2010, 2♂♂, 1♀ sobre Ulmus sp., 10.VII.2012, 1♂ sobre Q. rotundifolia, La Taha, Pitres, 23.VII.2010, 3♂♂, 3♀♀ sobre Castanea sativa. Zaragoza, Caspe, 110 m, 24.VI.2003, 1 ej., A. Gruppe (AG).
Coniopteryx (Holoconiopteryx) haematica McLachlan, 1868 (Figs. 189-194, 297, 298)
Coniopteryx haematica McLachlan, 1868
McLachlan, 1868: 193 (d); Monserrat, 1975: 264 (list); Marín & Monserrat, 1987: 350-359 (bio, dis); Monserrat & Marín, 1992: 283 (bio); Marín, 1994: 260-297 (bio, dis, fen); Monserrat & Triviño, 2013: 20 (list); Letardi et al., 2013: 36 (list).
Coniopteryx (Holoconiopteryx) haematica McLachlan, 1868
Monserrat, 1975: 264 (list, mf); Aspöck et al., 1980: 151 (bio, mf, tx, rf); Monserrat, 1981c: 179-182 (bio, dis, mf), 1982a: 83 (bio, dis), 1984a: 147 (dis, tx), 1984b: 107 (bio, dis), 1984c: 161 (rf), 1984d: 43 (bio,
dis), 1984e: 176 (bio, dis), 1985b: 89 (bio, dis), 1985d: 131 (bio, dis); Díaz-Aranda et al., 1986a: 1125 (bio, dis), 1986b: 1142 (bio, dis); Monserrat, 1987a: 142 (bio, dis); Monserrat & Díaz-Aranda, 1987: 183 (bio, dis); Díaz-Aranda & Monserrat, 1988a: 120 (bio, dis), 1988b: 227 (bio, dis), 1988c: 224 (bio, dis); Monserrat & Díaz-Aranda, 1988a: 93 (bio, dis), 1989: 54 (bio, dis); Marín & Monserrat, 1989: 296 (bio, dis); Monserrat & Díaz-Aranda, 1990: 43 (bio, dis); Marín & Monserrat, 1990: 228 (bio, dis), 1991b: 183 (bio, dis); Monserrat et al., 1994: 58 (bio, dis); Marín & Monserrat, 1995a: 44 (bio, dis), 1995b: 119 (bio, dis); Monserrat, 1995: 122 (bio, dis), 1996a: 15 (list); H. Aspöck et al., 2001: 187 (rf); Monserrat, 2002: 12 (bio, dis), 2005a: 74 (rf), 2013: 300 (bio, dis); Monserrat et al., 2013: 55 (bio, dis); Monserrat & Triviño, 2013: 20 (list), 120 (dis).
Coniopteryx sp.
Monserrat, 1977a: 171 (key, mf), 172 (bio, dis).
Coniopteryx (Holoconiopteryx) haematica fue brevemente descrita de Italia (sin más datos) por McLachlan (1868) en base a tres sintipos (♀♀) que fueron posteriormente
estudiados por Meinander (1972a), quien designa a uno de ellos lectotipo y propone como sinonimia de esta especie a Coniopteryx tullgreni Tjeder, 1930, especie que había sido descrita de Suecia y de la que ahora haremos referencia, y que desde entonces así ha sido mayoritariamente
considerada.
Coniopteryx (Holoconiopteryx) haematica es una especie citada como Holo-mediterránea y referenciada de Europa, en especial del centro y sur: Italia, Sicilia, Portugal,
España, Francia, Córcega, Croacia, Austria, Bulgaria, Hungría, Polonia, Rumanía, Suecia, Eslovenia, Rusia, Grecia, N. África
(Marruecos y Argelia), N. Turquía (Anatolia) y Chipre (Meinander, 1972a, 1981, 1990; Gepp, 1974b, 1977; Aspöck et al., 1980, 2001; Dorokhova, 1987, etc.). Posee una marcada preferencia por Quercus, pero también ha sido recolectada sobre Populus, Crataegus, Ceratonia, Pistacea, Citrus, Olea, Carpinus y Prunus, más escasamente sobre coníferas (Juniperus y Pinus) y ocasionalmente sobre Fragaria y emergiendo de agallas de Andricus kollari (Htg.), entre V-VIII, en altitudes de 100 hasta 1860 m. En nuestra fauna está ampliamante citada, sobre todo en zonas de
influencia mediterránea (incluido Baleares: Menorca), y ha sido citada sobre encinas, alcornoques, robles, quejigos, enebros,
pinos, chopos, arces, sauces, algarrobos, pistáceas, endrinos, frutales, a veces emergiendo de sus agallas, entre III-IX (los
datos ahora anotados en el nuevo material premiten sugerir actividad durante el invierno, tanto de larvas, como de imagos,
en la zona mediterránea), en altitudes que oscilan entre 75-1310 m.
Por tratarse de dos especies próximas (Figs. 179-188/ 189-194, 297, 298), probablemente ha sido confundida en ocasiones con C. drammonti Rousset, 1964 (Aspöck et al., 1980), pero por lo que ahora comentaremos, es aún más probable que muchos de los datos de Coniopteryx (Holoconiopteryx) haematica (según nuestra opinión limitada a la zona mediterránea) estén mezclados con los de Coniopteryx (Holoconiopteryx) tullgreni (según nuestra opinión limitada al centro y norte de Europa), especies hasta ahora consideradas sinónimas. Coniopteryx (Holoconiopteryx) tullgreni fue descrita y citada de Suecia (Tjeder, 1930, 1931, 1967; Kimmins, 1934; Meinander, 1962a), y también ha sido citada (ocasionalmente, por lapsus, dentro de otros géneros como Sconiopteryx sic.) de Francia (Rousset, 1960b, 1964), Alemania (Ohm & Remane, 1968; Aspöck & Aspöck, 1969; Lauterbach, 1970), Austria (Aspöck, 1963b; Aspöck & Aspöck, 1964, 1969; Ressl, 1964a, 1964b, 1971; Gepp, 1967; Hölzel, 1973), Bohemia (Zelený, 1962), Rumanía (Kis, 1964b, 1969, 1972, 1975; Kis et al., 1970), Hungría (Sziráki, 1992c); Grecia (Aspöck & Aspöck, 1969) y Anatolia (Aspöck & Aspöck, 1969). La genitalia femenina de esta especie fue descrita por Tjeder (1964) (Figs. 299-300).
Sinonimizadas ambas especies por Meinander (1972a), lo conocido de ambas especies se solapó, y en la mayoría de las citas posteriores (salvo las anteriormente anotadas) ya
solo se mencionaba a C. haematica, por lo que no podemos discernir si pudieran referirse a una u otra especie y por ello no los citamos (ver reseñas en Aspöck et al., 1980, 2001 o Meinander, 1990), y los datos de biología, fenología y distribución de una y otra especie se sumaron en los trabajos generales (Meinander, 1972a, 1990; Aspöck et al., 1980, 2001; Sziráki, 2004, 2011).
En base a los datos conocidos, ya Monserrat (1977a, 1981c) sostenía dudas sobre la identidad de los ejemplares ibéricos, por tener en la genitalia masculina el estilo bifurcado (Figs. 189-191), hecho que no ocurre (Fig. 188) en lo conocido para Coniopteryx (Holoconiopteryx) haematica (+ C. tullgreni sensu Tjeder, 1930) (Tjeder, 1930: 206, 1931: 24, lám. 4, fig.A, 1954: 27, fig. 5 B; Meinander, 1962a: 23, fig. 23, 1972a: 271, fig. 174A; Aspöck & Aspöck, 1964: 263, fig. 44; Rousset, 1964: 5, 8; Kis, 1964a: 52, fig. 12; Kis et al., 1970: 81, fig. 30 E; Aspöck et al., 1980: 58, fig. 291; Dorokhova, 1987: 45, fig, 3; Sziráki, 2004: 31, fig. 170A, 2011: 225, fig. 312A, etc.).
Este carácter (presencia de estilo bifurcado) (Figs. 189-191) ya fue detectado por Monserrat (1977a: 171-172) en los ejemplares ibéricos estudiados que, por precaución fueron inicialmente citados como Coniopteryx sp., pensando que pudiera tratarse de una nueva especie. Más tarde, y en base a nuevo material de España, Italia (continental)
y Rumanía, Monserrat (1981b, 1981c, 1995) asignó tentativamente este elemento como un carácter no previamente conocido en Coniopteryx (Holoconiopteryx) haematica McLachlan, 1868, y sugería un posible cierto grado de variabilidad entre poblaciones más meridionales respecto a las más septentrionales.
Posteriormente citaríamos abundante material ibérico con estas características (ver referencias anteriormente anotadas) y
también de Italia (Sicilia) o Marruecos (Monserrat, 1985e; Monserrat et al., 1991; Lo Verde & Monserrat, 1997), material ya publicado y que no consideramos necesario mencionar ahora en el nuevo material estudiado, y ahora solo anotamos
nuevo material de España, Portugal e Italia (Cerdeña).
El hecho es que todas las referencias existentes sobre la genitalia masculina de esta especie no coincidían con los ejemplares
estudiados por nosotros y todas ellas indicaban un estilo no bifurcado, corto, romo y robusto (Fig. 188), sin elementos de unión a los parámeros, y éstos con proceso dorsal simple, hechos que no ocurren en los ejemplares ibéricos,
italianos o rumanos estudiados por nosotros (Figs. 189-191), y aunque algunas descripciones de las genitalia masculina existentes en la bibliografía estaban (o parecen estar) basadas
en elementos previamente publicados (p. ej.: Meinander, 1962a; Dorokhova, 1987; Sziráki, 2004, 2011), era muy dudoso que ninguno de estos autores citados hubiera percibido la presencia de unos parámeros con proceso dorsal
más complejo (Fig. 193) y sobre todo de un estilo marcadamente bifurcado, con su rama caudal corta y de extremo agudo y su rama anterior fuertemente
esclerificada y arqueada que contacta con los parámeros (Figs.189-191).
Todos estos datos sugieren pues la existencia de dos especies próximas, pero diferentes: Coniopteryx (Holoconiopteryx) haematica McLachlan, 1868, en la zona europea más meridional, al sur de la barrera Pirineos-Alpes-Cárpatos-Cáucaso (constatada en la región mediterránea
occidental, y hemos estudiado abundante material de la Península Ibérica: España, incluido Baleares (Menorca), Portugal, Italia
continental, Sicilia (ahora citamos material de Cerdeña), Rumanía y Marruecos, y es lógico que a esta especie pertenezcan
las citas de Francia, y probablemente de zonas más orientales de Europa: Croacia, Hungría, Bulgaria, Rumanía, Eslovenia, Grecia,
Chipre y Anatolia, con estilo bifurcado en la genitalia masculina (Figs. 189-191), elemento no fácil de visualizar en ocasiones y que podría haber sido desapercibido por algunos autores que la citan de
estas zonas, datos que deberían corroborarse con el estudio del material de estos países, y por el contrario Coniopteryx (Holoconiopteryx) tullgreni Tjeder, 1930, del centro y norte de Europa, por encima de estas cadenas montañosas (Suecia, Alemania, Austria, República Checa, Polonia),
con estilo no bifurcado en la genitalia masculina (Fig. 188). También la genitalia interna femenina puede sugerir alguna diferencia (Figs. 297-300). Consecuentemente se propone revalidar a Coniopteryx (Holoconiopteryx) tullgreni Tjeder, 1930 como especie válida:
Coniopteryx (Holoconiopteryx) tullgreni Tjeder, 1930: 206 n. stat., sp. reval.
Que sepamos, los estadios preimaginales (de ambas especies) son desconocidos. Únicamente Monserrat et al. (1991: 108) hacen referencia, como probable, a la morfología del segundo estadio de C. haematica.
NUEVO MATERIAL Y MATERIAL COMPLEMENTARIO ESTUDIADO: ESPAÑA: Almería, Puerto de Santa María de Nieva, 1.085 m, 5.V.2007, 1♂, 1♀ en vuelo sobre Quercus rotundifolia. Ávila, Villaviciosa, 28.VII.2007, 1♂ sobre Q. rotundifolia. Islas Baleares, Menorca, Poblat de Torralba, 25.VII.2008, 1♂, 2♀♀ sobre Olea europaea, F. Acevedo, Torre d′en Galmés, 27.VII.2008, 4♂♂, 5♀♀ sobre O. europaea, F. Acevedo. Barcelona, Cabrils, 6.VIII.2000, 1♂, 2♀♀ a la luz, M. Carles-Tolrá. Cáceres, Almaraz, 39°47′16′′N, 5°41′59′′O, 315 m, 8.VII.2008, 1♀ sobre Q. rotundifolia. Cádiz, Pinsapar de Grazalema, 3.V.2005, 2♀♀ sobre Q. rotundifolia, 26.V.2012, 2♂♂, 1♀ sobre Q. canariensis, 22.VI.2012, 4♂♂, 2♀♀ sobre Q. rotundifolia, Puerto del Boyar, 22.VI.2012, 2♂♂ sobre Ceratonia siliqua. Castellón, Morella, 24.VIII.2006, 1♂, 1♀ sobre Q. rotundifolia. Ciudad Real, Alcoba, P. N. de Cabañeros, 1.II.2007, 1♂ sobre Q. rotundifolia. Granada, El Dornajo, 37°07’52.22’’N 3°26’57.43’’O, 1750 m, 11.VII.2010, 1♀ sobre Q. rotundifolia, La Herradura, Cerro Gordo, 6.VIII.2011, 1♂ a la luz, La Herradura, Punta de la Mona, 30SVF36, 200 m, 10.VIII.2010, 1♂ a
la luz. Jaén, Aldeaquemada, 38°23’44.86’’N 3°25’15.62’’O, 950 m, 24.VI.2009, 4♂♂ sobre Q. rotundifolia. Madrid, Fuentidueña de Tajo, 28.VI.2009, 1♀ sobre Q. rotundifolia. Málaga, Canillas de Albaida, Fábrica de la luz, Río de la Llanada de Turvilla, 30S 413487 4080230, 720 m, 28.VI.2015, 1♂, C. Zamora,
Puerto de Montejaque, 22.VI.2012, 1♀ sobre Q. rotundifolia. Salamanca, Berruecopardo, 31.V.2009, 1♀ sobre Q. pyrenaica. Tarragona, La Selva del Camp, 41°13′07.67′′N, 1°08′36.94′′E, 224 m (por su elevado número, solo anotamos parte del material estudiado)
27.XII.2005, 1♂, 1♀, 2 larvas, 17.VII.2010, 1♀, 17.VIII.2010, 1♂, 7.IV.2008, 2♂, 14.VII.2008, 1♀, 21.VIII.2008, 1♂, 18.XI.2008,
2♀♀, 23.IV.2010, 1♀, 16.VI.2010, 1♂, 10.VIII.2010, 1♂, 17.VIII.2010, 1♀, 11.VI.2011, 1♀, 16.VII.2011, 1♀, 15.X.2011, 1♀, todos
sobre Citrus clementina, N. Cañellas. ITALIA: Cerdeña: Cagliari, Iglesias, San Benedetto, 23.VIII.2005, 3♂♂, R. A. Pantaleoni. Nuoro, Neoneli, 40°03′57′′N, 8°56′56′′E, 545 m, 21.V.2008, 1♀ sobre Q. ilex, Lula, 15.VII.2005, 4♂♂ a la luz, C. Foxi. Oristano, Villa Verde, 31.VIII.2008, 6♂♂ a la luz, C. Foxi. Sassari, Alghero, Carrabuffas, 18.VIII.2005, 2♂♂, S. Cossu. PORTUGAL: Trás-os-Montes e Alto Douro, Bragança, 19.VI.2011, 5 ej. A. Gruppe (AG).
Coniopteryx (Metaconiopteryx) arcuata Kis, 1965 (Figs. 199, 200, 312)
Coniopteryx arcuata Kis, 1965
Kis, 1965b: 296 (d); Monserrat, 1977a: 22 (list); Monserrat & Marín, 1992: 283 (bio); Monserrat & Triviño, 2013: 20 (list).
Coniopteryx (Metaconiopteryx) arcuata Kis, 1965
Monserrat, 1977a: 174 (key, mf), 176 (bio, dis), 1978b: 183 (bio, dis), 1979c: 103 (bio, dis); Aspöck et al., 1980: 153 (bio, mf, tx, rf); Monserrat, 1982a: 84 (bio, dis), 1984a: 148 (dis, tx), 1984c: 161 (bio, dis), 1984d: 43 (bio, dis), 1984e: 175 (bio, dis); Díaz-Aranda et al., 1986b: 1143 (bio, dis); Monserrat & Díaz-Aranda, 1987: 184 (bio, dis); Marín & Monserrat, 1989: 296 (bio, dis), 1990: 228 (bio, dis); Monserrat & Díaz-Aranda, 1990: 43 (bio, dis); Monserrat et al., 1994: 59 (bio, dis); Marín & Monserrat, 1995b: 119 (bio, dis); Monserrat, 1996a: 15 (list); H. Aspöck et al., 2001: 189 (rf); Monserrat, 2002: 12 (bio, dis), 2005a: 74 (bio, dis), 2013: 301 (bio, dis); Monserrat et al., 2013: 55 (bio, dis); Monserrat & Triviño, 2013: 20 (list), 121 (dis).
Especie conocida de Europa, Marruecos y Turquía. Típica de medios relativamente húmedos de carácter mediterráneo, citada sobre
Quercus, Fagus, Crataegus, Alnus, Acer, Celtis, Corylus, Ulmus, Pyrus, Acacia, y Carpinus, mucho más escasamente sobre coníferas (Pinus), entre V-VIII, en altitudes de hasta 1350 m. En nuestra fauna ha sido ampliamente citada, incluyendo Baleares (Menorca),
recolectada sobre encinas, robles, quejigos, avellanos, castaños, alnueces, alisos, acacias o mimosas, entre V-IX, en altitudes
que oscilan entre 160-1320 m.
Que sepamos, los estadios preimaginales son desconocidos.
NUEVO MATERIAL Y MATERIAL COMPLEMENTARIO ESTUDIADO: ESPAÑA: Álava, Iruraitz-Gauna, Gazeo, 30TWN4644, 600 m, 8.VI.2007, 1♀ sobre Crataegus monogyna, S. Pagola, 4.VIII.2007, 1♀ sobre C. monogyna, S. Pagola. Ciudad Real, Navas de Estena, 16.V.2012, 1♂, 1♀ sobre Quercus faginea, 17.V.2012, 1♂ sobre Q. pyrenaica, F. Acevedo. Granada, Cáñar, 30SVF6389, 1354 m, 6.V.2011, 7♂♂, 7♀♀ sobre Castanea sativa. Salamanca, Berruecopardo, 31.V.2009, 2♀♀ sobre Q. pyrenaica, Gomeciego, 31.V.2009, 3♀♀ sobre Q. pyrenaica, El Milano, 31.V.2009, 1♂, 1♀ sobre Q. pyrenaica, Lumbrales, 31.V.2009, 1♂ sobre Q. pyrenaica. Tarragona, La Selva del Camp, 41°13′07.67′′N, 1°08′36.94′′E, 224 m, 17.VIII.2008, 1♀, 17.VIII.2010, 4♂♂, todos sobre Citrus clementina, N. Cañellas. ITALIA: Piemonte, Alessandria, Stazzano, 20.VIII.1985, 1♂, leg.? (MG). Sardegna, Nuoro, Fonni, 40°07′09′′N, 9°15′16′′E, 971 m, 21.V.2008, 2♂♂, 2♀♀ sobre Q. pubescens, 1♂, 1♀ sobre C. sativa, Neoneli, 40°03′57′′N, 8°56′56′′E, 545 m, 21.V.2008, 1♂ sobre Q. ilex.
Coniopteryx (Metaconiopteryx) esbenpeterseni Tjeder, 1930 (Figs. 195, 196, 315)
Coniopteryx esbenpeterseni Tjeder, 1930
Tjeder, 1930: 206 (d); Monserrat, 1977a: 22 (list), 1977c: 140 (bio, mf), 142 (list); Monserrat & Marín, 1992: 283 (bio); Ribeiro, 1997: 256 (bio, dis); Monserrat & Triviño, 2013: 20 (list); Letardi et al., 2013: 36 (list).
Coniopteryx (Metaconiopteryx) esbenpeterseni Tjeder, 1930
Monserrat, 1977a: 174 (key, mf), 177 (bio, dis), 1977c: 139 (rf), 140 (bio, dis, mf), 142 (rf), 1978b: 183 (bio, dis); Aspöck
et al., 1980: 152 (bio, mf, tx, rf); Castellari, 1980: 158 (rf); Monserrat, 1984a: 147 (dis, tx), 1984c: 161 (bio, dis), 1984d: 43 (bio, dis), 1984e: 175 (bio, dis), 1984g: 32 (bio, dis), 1985a: 95 (bio, dis); Díaz-Aranda et al., 1986b: 1143 (bio, dis); Monserrat & Díaz-Aranda, 1987: 183 (bio, dis); Marín & Monserrat, 1989: 296 (bio, dis); Monserrat & Díaz-Aranda, 1990: 43 (bio, dis); Monserrat et al., 1994: 58 (bio, dis); Marín & Monserrat, 1995b: 119 (bio, dis); Monserrat, 1995: 122 (bio, dis), 1996a: 15 (list); H. Aspöck et al., 2001: 189 (rf); Monserrat, 2002: 12 (dis, bio); Monserrat et al., 2013: 55 (bio, dis); Monserrat & Triviño, 2013: 20 (list), 121 (dis).
Elemento holomediterráneo extendido desde centroeuropa hasta Armenia. Está mayoritariamente citada sobre planifolios, entre
IV-IX, en altitudes de hasta 1700 m. En nuestra fauna ha sido generalmente citada en medios húmedos sobre robles, avellanos,
bojs, arces, abedules, acebos, sauces, coscoja, frutales, etc., pudiendo colonizar zonas más xéricas ocupando microhábitats
más húmedos sobre encinas, alnueces, algarrobos, tarajais, etc., recolectada entre III-IX, en altitudes que oscilan entre
30-1890 m.
Los estadios preimaginales fueron descritos por Castellari (1980: 168).
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Tarragona, La Selva del Camp, 41°13′07.67′′N, 1°08′36.94′′E, 224 m, 8.III.2008, 1♀, 21.VIII.2008, 1♂, 3♀♀, 15.IX.2008, 1♂, 17.VIII.2010,
1♀, 10.IV.2011, 1♀, 11.VI.2011, 2♂♂, todos sobre Citrus clementina, N. Cañellas.
Coniopteryx (Metaconiopteryx) lentiae Aspöck & Aspöck, 1964 (Figs. 197, 198, 314)
Coniopteryx lentiae Aspöck & Aspöck, 1964
Aspöck & Aspöck, 1964: 148 (d); Monserrat & Marín, 1992: 283 (bio); Marín, 1994: 260-297 (bio, dis, fen); Hynd, 1989: 160 (rf); Monserrat & Triviño, 2013: 20 (list).
Coniopteryx (Metaconiopteryx) lentiae Aspöck & Aspöck, 1964
Monserrat, 1977a: 177 (nt), 1978b: 183 (bio, dis); Aspöck et al., 1980: 153 (bio, mf, tx, rf); Monserrat, 1982a: 84 (bio, dis), 1984a: 148 (dis, tx), 1984b: 108 (bio, dis), 1984d: 43 (bio, dis), 1984e: 175 (bio, dis), 1984g: 32 (bio,
dis), 1985a: 95 (bio, dis); Díaz-Aranda et al., 1986a: 1125 (bio, dis), 1986b: 1143 (bio, dis); Marín & Monserrat, 1989: 296 (bio, dis), 1990: 228 (bio, dis), 1991b: 183 (bio, dis); Sziráki, 1992c: 359 (rf); Monserrat et al., 1994: 59 (bio, dis); Marín & Monserrat, 1995b: 119 (bio, dis); Monserrat, 1995: 123 (bio, dis), 1996a: 15 (list); H. Aspöck et al., 2001: 189 (rf); Monserrat et al., 2013: 55 (bio, dis); Monserrat & Triviño, 2013: 20 (list), 122 (dis).
Especie holomediterránea conocida de Europa central y meridional, Anatolia, Israel, Líbano e Irán. Muy euritópica y de tendencia
termófila. Generalmente habita en medios húmedos, pero también coloniza medios-microhabitats más húmedos en zonas más térmicas
y xéricas mediterráneas, y ha sido recolectada sobre vegetación arbórea, generalmente caducifolia (Corylus, Quercus, Ulmus, Castanea, Acer, Salix, Populus, Tamarix, Fagus, Carpinus, Genista, Evonymus, Crataegus, Tilia y Juglans), también sobre frutales y más escasamente sobre coníferas (Juniperus) y vegetación de rivera, entre V-VIII, en altitudes de hasta 1700 m. En nuestra fauna es conocida de los dos tercios más
septentrionales de la península, y ha sido citada sobre similares plantas, entre V-VIII, en altitudes que oscilan entre 220-1310
m.
Que sepamos, los estadios preimaginales son desconocidos.
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Ciudad Real, Navas de Estena, 17.V.2012, 1♂ sobre Quercus pyrenaica, F. Acevedo. Palencia, Palenzuela, 23.VIII-8.IX.2001, 1♂, 1♀ en trampa de Malaise, J. F. Gómez.
Coniopteryx (Metaconiopteryx) tjederi Kimmins, 1934 (Figs. 201, 202, 313)
Coniopteryx tjederi Kimmins, 1934
Kimmins, 1934: 613 (d); Monserrat, 1977a: 22 (list), 1977c: 140 (dis, mf), 142 (list), 1978a: 67 (bio, dis); Monserrat & Marín, 1992: 283 (bio); Marín, 1994: 260-297 (bio, dis, fen); Monserrat & Triviño, 2013: 20 (list); Letardi et al., 2013: 36 (list).
Coniopteryx (Metaconiopteryx) tjederi Kimmins, 1934
Monserrat, 1977a: 173 (key, mf), 175 (bio, dis, mf), 1977c: 139 (rf), 140 (bio, dis, mf), 142 (list), 1980: 192 (bio, dis);
Aspöck et al., 1980: 154 (bio, mf, tx, rf); Meinander, 1981: 107 (dis); Monserrat, 1982a: 84 (bio, dis), 1984a: 148 (dis, tx), 1984d: 43 (bio, dis), 1985a: 95 (bio, dis), 1985b: 90 (bio, dis), 1985d: 131 (bio, dis); Díaz-Aranda et al., 1986a: 1125 (bio, dis), 1986b: 1143 (bio, dis); Monserrat & Díaz-Aranda, 1987: 184 (bio, dis), 1988a: 93 (bio, dis); Díaz-Aranda & Monserrat, 1988a: 121 (bio, dis), 1988b: 227 (dis); Monserrat & Díaz-Aranda, 1989: 54 (bio, dis), 1990: 43 (bio, dis); Marín & Monserrat, 1990: 227 (bio, dis), 1991b: 183 (bio, dis); Monserrat et al., 1994: 59 (bio, dis); Marín & Monserrat, 1995a: 44 (bio, dis), 1995b: 119 (bio, dis); Monserrat, 1995: 122 (bio, dis), 1996a: 15 (list); H. Aspöck et al., 2001: 190 (rf); Monserrat, 2002: 12 (bio, dis), 2013: 301 (bio, dis); Monserrat & Triviño, 2013: 20 (list), 122 (dis).
Especie holomediterránea conocida de Europa central y meridional, Marruecos, Asia Menor y Ucrania. Muy euritópica y citada
generalmente sobre vegetación arbórea caducifolia (Corylus, Quercus, Ulmus, Prunus, Salix, Populus, Alnus, Fraxinus, Arundo, Sarothamnus y Juglans), también sobre frutales, vegetación de rivera, leguminosas y más escasamente sobre coníferas (Pinus y Juniperus). También coloniza zonas mucho más xéricas en hábitats-microhabitats más húmedos sobre fanerófitos mediterráneos (Ceratonia, Quercus, Celtis, Ulmus, Pistacea, Arundo, Tamarix o Crataegus) y vegetación de rivera, escasamente sobre Pinus. Citada entre III-VII, en altitudes de hasta 2000 m. En nuestra fauna ha sido citada sobre estas plantas, entre V-IX, en
altitudes que oscilan entre 10-1480 m.
Que sepamos, los estadios preimaginales son desconocidos.
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Cádiz, Pinsapar de Grazalema, 3.V.2005 1♂, 1♀, sobre Quercus canariensis. Granada, Cáñar, 30SVF6389, 1354 m, 6.V.2011, 1♂ sobre Castanea sativa, 27.VI.2011, 2♀♀ sobre C. sativa, 3♀♀ sobre Q. pyrenaica, 18.VII.2011, 2♀♀ sobre Q. pyrenaica, La Herradura, Punta de la Mona, 30SVF36, 200 m, 24.IV.2011, 1♂ sobre Acacia dealbata, La Taha, Pitres, 23.VII.2010, 2♂♂ sobre C. sativa. Huelva, Castaño de Robledo, 23.V.2012, 1♂, 1♀ sobre Q. canariensis. Madrid, El Escorial, Arboreto Luis Ceballos, 1.VIII-12.IX.2008, 1♂ en trampa de Malaise. Zaragoza, Caspe, 110 m, 24.VI.2003, 1 ej., A. Gruppe (AG).
Coniopteryx (Xeroconiopteryx) atlantica Ohm, 1963 (Figs. 203-210)
Coniopteryx atlantica Ohm, 1963
Ohm, 1963: 267 (d); Monserrat & Marín, 1992: 282 (bio); Monserrat & Triviño, 2013: 20 (list).
Coniopteryx (Xeroconiopteryx) atlantica Ohm, 1963
Monserrat, 1985d: 128 (rf), 131 (dis, mf), 1987a: 143 (bio, dis), 145 (rf); Monserrat & Díaz-Aranda, 1990: 42 (dis); Meinander, 1990: 42 (rf); Monserrat, 1995: 115 (dis); H. Aspöck et al., 2001: 180 (rf); Sziráki, 2004: 34 (rf); Monserrat, 2013: 301 (rf); Monserrat & Triviño, 2013: 20 (list), 123 (dis).
Especie conocida del S.E. de España, Islas Canarias (Tenerife, Fuerteventura, ahora citada de La Palma), República Árabe Sahariana
Democrática y Marruecos. Muy escasamente citada, usualmente recolectada a la luz en zonas xéricas y en hábitats térmicos con
halófitas y Chenopodiaceae, a veces en medios urbanos, generalmente en las proximidades del mar, algunos imagos se han recolectado
sobre Juniperus. Ha sido generalmente citada en I-V, en altitudes bajas y medias, ahora la citamos (VII, IX) hasta 1068 m de La Palma. En
la fauna tratada ha sido citada en similares medios, recolectada a la luz en ramblas subdesérticas (Almería), entre V-VII,
en altitudes que oscilan entre 10-400 m.
Que sepamos, los estadios preimaginales son desconocidos.
NUEVO MATERIAL COMPLEMENTARIO ESTUDIADO: ESPAÑA: Islas Canarias, La Palma, P. N. de La Caldera de Taburiente, Barranco de las Traves, 1068 m, 28RBS1780-2, 24.VII.2000, 1♀ en trampa de Malaise, T.
Domingo (MNCN), 18.IX.2000, 1♀ en trampa de Malaise, T. Domingo, Playa de Taburiente, 750 m, 28RBS1980-1, 5.IX.2000, 1♀ en
trampa de Malaise, T. Domingo.
Coniopteryx (Xeroconiopteryx) atlasensis Meinander, 1963 (Figs. 18, 241-242)
Coniopteryx atlasensis Meinander, 1963
Meinander, 1963:101 (d); Monserrat, 1977a: 22 (list); Monserrat & Marín, 1992: 282 (bio); Marín, 1994: 260-297 (bio, dis, fen); Monserrat, 2010: 28 (rf); Monserrat & Triviño, 2013: 20 (list).
Coniopteryx (Xeroconiopteryx) atlasensis Meinander, 1963
Monserrat, 1976b: 247 (rf), 249 (bio, dis), 1977a: 163 (key, mf), 165 (nt), 166 (bio, dis), 1980: 192 (bio, dis); Aspöck et al., 1980: 154 (bio, mf, tx, rf); Monserrat, 1982a: 83 (bio, dis), 1982b: 61 (rf), 1984a: 147 (dis, tx), 1984b: 107 (bio, dis), 1984e: 176 (bio, dis), 1984g: 32 (bio, dis),
1985b: 89 (bio, dis); Díaz-Aranda et al., 1986b: 1143 (bio, dis); Monserrat, 1987a: 143 (bio, dis); Monserrat & Díaz-Aranda, 1987: 183 (bio, dis), 1988a: 93 (bio, dis); Díaz-Aranda & Monserrat, 1988a: 121 (dis), 1988c: 224 (bio, dis); Monserrat & Díaz-Aranda, 1989: 54 (bio, dis), 1990: 42 (bio, dis); Meinander, 1990: 42 (rf); Marín & Monserrat, 1990: 228 (bio, dis), 1991b: 183 (bio, dis); Monserrat et al., 1994: 57 (bio, dis); Marín & Monserrat, 1995a: 43 (bio, dis), 1995b: 118 (bio, dis); Monserrat, 1995: 115 (bio, dis), 1996a: 14 (list); H. Aspöck et al., 2001: 181 (rf); Monserrat, 2002: 8 (bio, dis, mf); Sziráki, 2004: 37 (rf); Sziráki & Mirmoayedi, 2012: 47 (rf); Monserrat, 2013: 301 (rf); Monserrat & Triviño, 2013: 20 (list), 123 (dis).
Coniopteryx atlasiensis Meinander, 1963 (lapsus)
Gruppe, 2007b: 141 (rf).
Especie citada del límite meridional de la Región Paleártica occidental: Europa (España, Portugal, Creta), Marruecos, Islas
Canarias (Fuerteventura, La Palma), Madeira, Turquía, Yemen y Asia central (Afghanistán, Irán, Tadshikistán, Kazakhstán, Aserbeidshán).
Está mayoritariamente citada en medios térmicos y xerófilos de hábitats subdesérticos y mediterráneos. En Europa la mayor
parte de sus imagos se han recolectado a la luz, pero también ocasionalmente sobre Pinus, Quercus, Ceratonia, Tamarix, Juniperus, Populus, Ulmus, Pistacea y Olea. En África y Asia sobre Tamarix, Juniperus, Pinus, Quercus, Tetraclinis, Olea y Ceratonia. Probablemente esta especie se desarrolle sobre vegetación baja de carácter xerófilo. Está citada entre II-IX, en altitudes
de hasta 2180 m. En nuestra fauna ha sido recolectada en toda la mitad sud-oriental, principalmente a la luz y citada sobre
encinas, quejigos, coscojas, algarrobos, pinos, enebros, olivos, almendros, pistáceas, tarajais, chopos, olmos, entre III-IX,
en altitudes medias y bajas que oscilan entre 10-960 m. Ahora se amplían significativamente estos datos.
Que sepamos, los estadios preimaginales son desconocidos.
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Badajoz, Santa Marta de los Barros, 19.VI.2012, 1♂, 1♀ sobre Quercus rotundifolia. Cádiz, Algodonales, 26.V.2012, 1♂, 1♀ sobre Pistacea lentiscus. Granada, Ferreira, 30SVG9513, 1356 m, 8.V.2011, 2♂♂, 1♀ sobre Q. rotundifolia, Lobras, 30SVF8084, 822 m, 6.V.2011, 3♂♂, 1♀ sobre Q. rotundifolia. Huelva, La Nava, 23.V.2012, 1♀ sobre Q. rotundifolia. Madrid, Madrid, Ciudad Universitaria, 19.V.2010, 1♂ errante (ejemplar extraviado), Murcia, Rambla de Portús, Canteras, 31.VIII.2005, 4♀♀ a la luz (probablemente estos ejemplares pertenezcan a la especie que tratamos).
Coniopteryx (Xeroconiopteryx) kerzhneri Meinander, 1971 (Figs. 16, 211-216, 291)
Coniopteryx (Aspoeckiana) kerzhneri Meinander, 1971
Meinander, 1971: 42 (d).
Coniopteryx (Xeroconiopteryx) kerzhneri Meinander, 1971
Meinander, 1981; 104 (rf); Monserrat, 1982b: 57 (bio, dis), 61 (list), 1985b: 90 (bio, dis), 1985d: 134 (bio, dis), 1987a: 143 (bio, dis); Meinander, 1990: 41 (rf); Monserrat & Díaz-Aranda, 1990: 42 (dis); H. Aspöck et al., 2001: 179 (rf); Monserrat, 2002: 10 (bio, dis, mf); Sziráki, 2004: 33 (rf); Sziráki & Harten, 2006: 394 (rf); Sziráki & Mirmoayedi, 2012: 47 (rf); Monserrat, 2013: 301 (rf), 305 (rf); Monserrat & Triviño, 2013: 20 (list), 124 (dis).
Coniopteryx kerzhneri Meinander, 1971
Monserrat, 1983b: 126 (rf), Monserrat & Marín, 1992: 282 (bio); Monserrat, 2010: 28 (rf); Monserrat & Triviño, 2013: 20 (list).
Elemento eremial distribuido por las zonas secas del Paleártico meridional y extendido desde Mongolia, Yemen, Irán y norte
de África (Egipto, Argelia) hasta la Península Ibérica. De biología prácticamente desconocida, ha sido mayoritariamente recolectada
a la luz, entre VI-IX, en altitudes de hasta 1130 m. La hembra citada como Coniopteryx (Xeroconiopteryx) sp. por Monserrat et al. (1991: 108) de Marruecos pertenece a esta especie (Fig. 291), adjudicando así algún nuevo dato para esta especie (V, 1210 m, sobre Atriplex halinus). En nuestra fauna también ha sido generalmente recolectada a la luz en medios extremadamente secos y térmicos del sudeste
peninsular (Murcia, Almería y Granada), también sobre Tamarix, Pinus, Arundo, Ceratonia, entre V-X (probablemente sus poblaciones sufran diapausa estival), en altitudes bajas que oscilan entre 10-500 m.
Que sepamos, los estadios preimaginales son desconocidos.
Coniopteryx (Xeroconiopteryx) ketiae Monserrat, 1985 (Figs. 217-223)
Monserrat, 1985d: 134 (d); Monserrat & Díaz-Aranda, 1990: 42 (bio, dis); Meinander, 1990: 41 (rf); Monserrat, 1995: 116 (dis); Sziráki, 1997: 278 (rf); H. Aspöck et al., 2001: 180 (rf); Sziráki, 2004: 37 (rf); Monserrat, 2006: 19 (rf); Sziráki & Harten, 2006: 407 (rf); Monserrat, 2013: 301 (rf); Monserrat & Triviño, 2013: 20 (list), 124 (dis).
Coniopteryx ketiae Monserrat, 1985
Monserrat & Marín, 1992: 282 (bio); Monserrat, 2010: 28 (rf); Monserrat & Triviño, 2013: 20 (list).
Elemento inicialmente ibérico, del que ha sido extendida su distribución a otras zonas subdesérticas del Paleártico occidental
(Senegal, Yemen). Ha sido mayoritariamente recolectada a la luz, también sobre Arundo y plantas halófilas y xerófilas, entre II-X, en altitudes de 10 hasta 2253 m. En nuestra fauna ha sido mayoritariamente recolectada
a la luz en medios extremadamente secos y térmicos del S.E. peninsular (Murcia, Almería), algún ejemplar ha sido recolectado
sobre vegetación herbácea o sobre Arundo donax, entre VII-X (probablemente sus poblaciones sufran diapausa estival), en altitudes que oscilan entre 10-110 m.
Que sepamos, los estadios preimaginales son desconocidos.
Coniopteryx (Xeroconiopteryx) loipetsederi Aspöck, 1963 (Figs. 224-228, 306)
Coniopteryx loipetsederi Aspöck, 1963
Aspöck, 1963a: 95 (d); Aspöck & Aspöck, 1965a: 82 (rf); Kis, 1969: 37 (rf), 39 (rf); Zelený, 1971: 154 (rf); Monserrat, 1977a: 22 (list), 1977c: 142 (list), 1978a: 66 (bio, dis); Monserrat & Marín, 1992: 282 (bio); Marín, 1994: 260-297 (bio, dis, fen); Monserrat et al., 1999: 40, 42 (bio, dis); Monserrat & Triviño, 2013: 20 (list); Letardi et al., 2013: 36 (list).
Coniopteryx (Xeroconiopteryx) loipetsederi Aspöck, 1963
Meinander, 1972a: 224 (mf, rf); Monserrat, 1977a: 162 (key, mf), 164 (bio, dis); Aspöck et al., 1980: 155 (bio, mf, tx, rf); Monserrat, 1980: 192 (bio, dis), 1982a: 83 (bio, dis), 1982b: 61 (rf), 1984b: 107 (bio, dis), 1984d: 43 (bio, dis), 1984g: 32 (bio, dis),
1985b: 89 (bio, dis), 1985d: 131 (bio, dis), 1987a: 143 (bio, dis); Díaz-Aranda & Monserrat, 1988a: 121 (dis), 1988b: 227 (bio, dis), 1988c: 224 (dis); Monserrat & Díaz-Aranda, 1988a: 93 (bio, dis), 1989: 55 (bio, dis), 1990: 42 (bio, dis); Marín & Monserrat, 1991b: 183 (bio, dis); Monserrat et al., 1994: 57 (bio, dis); Marín & Monserrat, 1995a: 44 (bio, dis); Monserrat, 1995: 115 (bio, dis), 1996a: 15 (list); H. Aspöck et al., 2001: 180 (rf); Monserrat, 2002: 10 (bio, dis); Alvis et al., 2003: 186 (bio, dis); Monserrat, 2013: 301 (bio, dis); Monserrat et al., 2013: 55 (bio, dis); Canbulat, 2013: 243 (rf); Monserrat & Triviño, 2013: 20 (list), 125 (dis).
Coniopteryx (Aspoeckiana) loipetsederi Aspöck, 1963
Kis et al., 1970: 85 (rf).
Especie conocida del sur de Europa y Argelia, extendida hasta Anatolia e Irán, asociada a zonas térmicas y en ocasiones también
a zonas secas de medios mediterráneos. Sus imagos se han recolectado a la luz y sobre muy diferentes fanerófitos: (Olea, Tamarix, Juniperus, Ceratonia, Quercus, Pistacea, Myrtus, Acacia, Populus, Ulmus, Citrus y Pinus), aunque más frecuentemente lo ha sido a la luz, hecho que parece evidenciar su asociación con otro tipo de vegetación, probablemente
baja (generalmente mucho menos muestreada). Ha sido recolectada entre V-XI, en altitudes de 10 hasta 950 m. En nuestra fauna
ha sido mayoritariamente recolectada en los dos tercios sud-orientales de la península, principalmente a la luz, pero también
citada sobre álamos, acacias, cítricos, pinos, enebros, sabinas, encinas, olmos, pistáceas, algarrobos, olivos, tamariscos,
mirtos, alnueces, etc., también en espacios urbanos, entre IV-XI (probablemente posea actividad durante el invierno en zonas
mediterráneas), y ha sido recolectada en altitudes que oscilan entre 10-1480 m.
Que sepamos, los estadios preimaginales son desconocidos.
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Almería, Rambla del Retamar, 14.VI.2012, 1♀ a la luz, V. Triviño, Tabernas, Rambla Roja, 30SWG50, 400 m, 13.VI.2012, 1♀ a la luz,
F. Acevedo, 12.VII.2014, 3♀♀ sobre Tamarix sp., F. Acevedo. Ávila, Nava de Arévalo, 22.VIII-7.IX.2001, 2♂♂, 4♀♀ en trampa de Malaise, J. F. Gómez. Granada, Cádiar, 36°56’46’’N, 3°10’41’’O, 923 m, 5.VII.2010, 1♂ sobre Quercus rotundifolia, Cerro Gordo, 21.IV.2011, 1♀ sobre Pistacea lentiscus, La Herradura, 22.VI.2003, 3♂♂ a la luz. Murcia, Cabo de Palos, 4.IV.2004, 1♂ a la luz. Palencia, Palenzuela, 23.VIII-8.IX.2001, 3♂♂, 1♀ en trampa de Malaise, J. F. Gómez, 8-22.IX.2001, 1♂ en trampa de Malaise, J. F. Gómez.
Segovia, Chañe, 1-16.VIII.2000, 1♀ en trampa de Malaise, J. F. Gómez, 16.VIII-4.IX.2000, 1♂ en trampa de Malaise, J. F. Gómez.
Coniopteryx (Xeroconiopteryx) perisi Monserrat, 1976 (Figs. 15, 229-240)
Monserrat, 1976a: 97 (d), 1977a: 22 (list), 163 (key, mf), 165 (bio, dis); Meinander, 1979b: 337 (rf); Aspöck et al., 1980: 9 (dis), 155 (bio, mf, tx, rf); Meinander, 1981; 104 (rf); Monserrat, 1982a: 83 (bio, dis), 1982b: 61 (rf), 1984g: 32 (bio, dis), 1985b: 90 (bio, dis); Meinander, 1990: 43 (rf); Monserrat et al., 1991: 108 (rf); Marín & Monserrat, 1995b: 118 (bio, dis); Monserrat, 1995: 115 (bio, dis), 1996a: 15 (list); H. Aspöck et al., 2001: 181 (rf); Monserrat, 2002: 11 (bio, dis, mf) ; Sziráki, 2004: 37 (rf); Monserrat, 2013: 302 (bio, dis) ; Monserrat & Triviño, 2013: 20 (list), 125 (dis).
Coniopteryx perisi Monserrat, 1976
Monserrat, 1977a: 22 (list), 1977c: 142 (list); Monserrat & Marín, 1992: 282 (bio); Monserrat & Triviño, 2013: 20 (list).
Elemento conocido de la Península Ibérica y Marruecos. Está mayoritariamente citado en medios térmicos con escasa vegetación,
habiéndose recolectado sobre Pistacea, Pinus, Juniperus, Crataegus, Rosmarinus y Quercus, entre V-VIII, en altitudes de hasta 2200 m. En nuestra fauna ha sido recolectada en zonas interiores, mayoritariamente a
la luz y citada sobre encinas, endrinos, pinos, también en sabinares, entre IV-V, VII-IX (se deducen dos generaciones anuales),
en altitudes que oscilan entre 470-978 m.
Que sepamos, los estadios preimaginales son desconocidos.
Conwentzia pineticola Enderlein, 1905 (Figs. 263, 266)
Enderlein, 1905a: 10 (d); Navás, 1908: 128 (rf), 1910a: 54 (dis), 1910b: 244 (dis), 1913a: 85 (dis), 1914d: 190 (dis), 1915: 30 (dis), 1924a: 240, 241 (dis, key, mf), 1924b: 26 (dis); 1925a: 121
(dis, key, mf); Stitz, 1927: 18 (rf); Eglin, 1940: 269 (rf); Berland, 1962: 26 (rf); Monserrat, 1977a: 23 (list), 178 (key), 179 (bio, dis); Aspöck et al., 1980: 163 (bio, mf, tx, rf); Monserrat, 1984a: 148 (dis, tx), 1984c: 161 (bio, dis), 1984d: 44 (bio, dis), 1985a: 96 (bio, dis), 1985b: 91 (bio, dis), 1985d: 137 (bio, dis); Díaz-Aranda et al., 1986a: 1125 (bio, dis), 1986b: 1144 (bio, dis); Monserrat, 1987a: 144 (bio, dis); Séméria & Berland, 1988: 53 (rf); Díaz-Aranda & Monserrat, 1988b: 228 (bio, dis), 1988c: 225 (bio, dis); Monserrat & Díaz-Aranda, 1990: 46 (bio, dis); Monserrat & Marín, 1992: 283 (bio); Monserrat et al., 1994: 60 (rf); Marín & Monserrat, 1995a: 45 (bio, dis), 1995b: 120 (bio, dis); Monserrat, 1995: 124 (bio, dis), 1996a: 15 (list); H. Aspöck et al., 2001: 193 (rf); Monserrat, 2002: 14 (bio, dis), 2005a: 73 (bio, dis); Letardi, 2012: 71 (dis); Monserrat, 2013: 302 (bio, dis); Monserrat & Triviño, 2013: 20 (list), 126 (dis); Monserrat et al., 2013: 55 (bio, dis); Letardi et al., 2013: 36 (list).
Conventzia pineticola Enderlein, 1905 (lapsus)
Navás, 1919b: 43 (dis); Monserrat & Triviño, 2013: 20 (list).
Conventzia pineticola tetensi Klapálek, 1909 (lapsus)
Klapálek, 1909: 30 (dis); Monserrat & Triviño, 2013: 20 (list).
Durante un cierto tiempo esta especie ha sido considerada mera variabilidad de C. psociformis (Withycombe, 1922a; Lacroix, 1924; Killington, 1936, 1937b; Rousset, 1960b, etc.) por lo que las citas antiguas de ambas especies pueden estar mezcladas. Con nuevos elementos Collyer (1951) y Zelený (1961b) inician la correcta diferenciación de ambas especies.
Elemento holártico (probablemente extendido por acción humana a Canadá y USA). Especie mayoritariamente citada sobre coníferas,
entre IV-X, en altitudes de hasta 2000 m. En nuestra fauna ha sido ampliamente citada, incluyendo Baleares (Mallorca, Formentera),
sobre coníferas (Abies, Cedrus, Picea, Pinus, etc.), entre III-IX en altitudes que oscilan entre 10-2500 m. El ejemplar ahora citado, recolectado a 2800 m, mostraba la
membrana alar curiosamente oscura.
Los estadios preimaginales de esta especie (bajo diferentes denominaciones) fueron descritos por Quayle (1912: 506, 1913: 87), Withycombre (1923: 581), Collyer (1951: 558), Schremmer (1959: 122, 124), Rousset (1960b: 30), Principi & Canard (1974: 161).
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Granada, La Herradura, Punta de la Mona, 30SVF36, 200 m, 26.VIII.2010, 1♀ a la luz. Huelva, Matalascañas, 20.VI.2012, 1♀ sobre Juniperus phoenicea, Punta Umbría 20.VI.2012, 1♀ sobre Pinus pinea. Murcia, Cabo de Palos, 4.IV.2004, 3♀♀ a la luz. Segovia, Valsaín, 2.800 m, 8.VII.2007, 1♂ sobre P. sylvestris. PORTUGAL: Trás-os-Montes e Alto Douro, Vila Real, Sa. Do Marão, 840 m, 18.VI.2011, 1 ej., A. Gruppe (AG).
Conwentzia psociformis (Curtis, 1834) (Figs. 25, 41, 262, 264, 265, 307)
Coniopteryx psociformis Curtis, 1834
Curtis, 1834: 528 (d).
Conwentzia psociformis (Curtis, 1834)
Navás, 1908: 128 (dis, mf), 1910a: 40, 45, 46, 54 (dis), 1910b: 244 (dis), 1913a: 85 (dis), 1914d: 190 (dis), 1915: 30 (dis), 1916a: 175 (dis), 1916c: 190 (dis); Bohigas & Sanchez, 1917: 317 (dis); Navás, 1921a: 152 (dis), 1923b: 8 (dis), 1924a: 240 (dis, key, mf), 1924b: 26 (dis), 1925a: 120, 121 (dis, key, mf); Stitz, 1927: 18 (rf); Navás, 1929a: 33 (dis), 1931: 121 (dis); Esben-Petersen, 1931: 5 (dis); Killington, 1936: 193 (rf); Eglin, 1940: 268 (rf); Vidal y López, 1943: 24 (dis); Berland, 1962: 25 (rf); Hölzel, 1964: 111 (rf); Meinander, 1976: 88 (rf); Monserrat, 1977a: 23 (list), 178 (key, rf), 1978b: 182 (bio, dis), 1979a: 414 (bio, dis); Aspöck et al., 1980: 162 (bio, mf, tx, rf); Ripolles & Melia, 1980: 61 (bio, dis, h, l, p); Monserrat, 1980: 192 (bio, dis), 1982a: 82 (bio, dis); García Marí et al., 1983: 196, 200 (bio, dis, l, p); Monserrat, 1984a: 148 (dis, tx), 1984b: 109 (bio, dis), 1984d: 44 (bio, dis), 1984e: 176 (bio, dis), 1984g: 33 (bio, dis), 1985a: 96 (bio, dis), 1985d: 137 (bio, dis); Díaz-Aranda et al., 1986a: 1125 (bio, dis), 1986b: 1144 (bio, dis); Monserrat, 1987a: 144 (bio, dis); Marín & Monserrat, 1987: 350-359 (bio, dis); Monserrat & Díaz-Aranda, 1987: 185 (bio, dis), 1988a: 94 (bio, dis); Séméria & Berland, 1988: 51, 52 (rf); Díaz-Aranda & Monserrat, 1988a: 121 (bio, dis), 1988b: 228 (bio, dis), 1988c: 225 (bio, dis); Ros et al., 1988: 7, 8, 9 (bio, dis); Marín & Monserrat, 1989: 296 (bio, dis); Monserrat & Díaz-Aranda, 1989: 55 (dis), 1990: 46 (bio, dis); Marín & Monserrat, 1991b: 183 (bio, dis); Monserrat & Marín, 1992: 283 (bio); Marín, 1994: 260-297 (bio, dis, fen); Monserrat et al., 1994: 60 (rf); Marín & Monserrat, 1995a: 45 (bio, dis), 1995b: 120 (bio, dis); Monserrat, 1995: 124 (bio, dis), 1996a: 15 (list); Lo Verde & Monserrat, 1997: 62 (rf); García Marí & González Zamora, 1999: 490 (bio, dis); H. Aspöck et al., 2001: 193 (rf); Monserrat, 2002: 14 (bio, dis); Soler et al., 2002: 135, 142 (bio, dis, l); Alvis et al., 2003: 186 (bio, dis); Monserrat, 2005a: 73 (bio, dis); Ocharan et al., 2012: 246 (rf); Letardi, 2012: 72 (dis); Monserrat, 2013: 302 (rf); Monserrat & Triviño, 2013: 20 (list), 126 (dis); Monserrat et al., 2013: 55 (bio, dis); Letardi et al., 2013: 36 (list).
Conventzia psociformis (Curtis, 1834) (lapsus)
Navás, 1919b: 43 (dis); Monserrat & Triviño, 2013: 20 (list).
Especia paleártica (¿holártica?) extendida por acción humana a Canadá, USA y Nueva Zelanda. Está mayoritariamente citada sobre
una enorme variedad de plantas, especialmente planifolios, entre V-X, en altitudes de hasta 1920 m. Aunque se había dudado
de su presencia en la Península Ibérica, quizás poniendo en tela de juicio de las citas previas existentes (Meinander, 1976: 88), en nuestra fauna ha sido posteriormente citada con frecuencia sobre una enorme variedad de vegetales, mayoritariamente
planifolios, incluso frutales y ornamentales, muy excepcionalmente sobre coníferas, entre IV-X, en altitudes que oscilan entre
10-1400 m. Aunque en nuestra fauna parece tener una tendencia eurosiberiana (bojs, robles, avellanos, acebos, hiedras, etc.),
ha sabido ocupar otros nichos y micro-hábitats colonizando zonas marcadamente mediterráneas (encinas, alcornoques, madroños,
pistáceas, algarrobos, olivos), incluyendo Baleares (Mallorca, Menorca). A partir del material ahora citado y el recolectado
seriadamente por el Instituto Andaluz de Investigación y Formación Agraria (mediante trampas de intersección y de caída colgadas
en dos cultivos de aguacate en la provincia de Málaga), y por la Universidad Autónoma de Barcelona (mediante diferentes métodos
de muestreos en una parcela de cítricos en la provincia de Tarragona), se demuestra su actividad de imagos y de larvas en
medios naturales y en estos tipos de cultivos en la zona mediterránea de España a lo largo de todo el año, con sucesión de
generaciones que sortean los meses del invierno en actividad, tanto en estado de larva como de imago, y Monserrat (2002) anota datos de XI-XII en esta zona levantina sobre cítricos. Estos datos se corroboran con estudios realizados por García Marí & González Zamora (1999), Soler et al. (2002) y Alvis et al. (2003) en cultivos de cítricos en Castellón y Valencia. Monserrat (1995) cita también abundante material en Setúbal recolectado en II-V.
Los estadios preimaginales de esta especie (Figs. 25, 41) fueron descritos o reiterados por Schlechtendal (1882: 26, 1883: 70), Enderlein (1906: 185); Arrow (1917: 254), Withycombe (1922b: 224; 1923: 579; 1924b: 115; 1925a: 410), Lacroix (1924: 70), Withycombre (1925a: 319), Stitz (1931: 101), Killington (1936: 190), Collyer (1951: 559), Obenberger (1958: 457); Rousset (1960b: 30), Zelený (1961a: 170), Ghilarov (1962: 412), Hoffmann (1962: 323), Rousset (1966: 53, 1969: 111): Kis et al. (1970: 58); Principi & Canard (1974: 161); Ripolles & Melia (1980: 61); García Marí et al. (1983: 196); Gepp (1990: 133, 1999: 194); Sinacori et al. (1992: 29), Soler et al. (2002: 142). Citada sobre Viburnum tinus (Adoxaceae) y erróneamente asignada a Hemerobius hirtus, fue descrita por Dujardin (1851).
NUEVO MATERIAL Y MATERIAL COMPLEMENTARIO ESTUDIADO: EGIPTO: Giza, Atfieh, VIII.2005, 1♂ sobre Brassica oleracea, infestada por agromízidos minadores de sus hojas, S.A.El-Serwy (VM). ESPAÑA: Almería, El Molino de La Serena, Bédar, 1.V.2007, 1♀ a la luz, 10♂♂, 9♀♀ sobre Citrus sp., 17.VII.2007, 1♂, 1♀ sobre Pistacea lentiscus, 1♂ sobre Ceratonia siliqua, La Serena, Bédar, 1.V.2007, 1♂, 2♀♀ sobre C. siliqua, 1♂, 1♀ sobre Rhamnus alaternus. Ávila, Embalse de Burguillo, Barraco, 12.V.2007, 1♀ sobre Hedera helix. Islas Baleares, Menorca, Poblat de Torralba, 25.VII.2008, 2♂♂, 3♀♀ sobre Olea europaea, F. Acevedo. Torre d′en Galmés, 27.VII.2008, 1♂, 2♀♀ sobre O. europaea, F. Acevedo. Barcelona, Cabrils, 6.VIII.2000, 2♀♀ a la luz, M. Carles-Tolrá. Cádiz, Barbate, Caños de Meca, 20.VI.2012, 2♂♂ sobre P. lentiscus, 1♂ sobre O. europea, Facinas, 25.V.2012, 1♂ sobre O. europea, de Grazalema, 3.V.2005, 1♀ sobre Quercus canariensis, 1♂ sobre Q. suber, 3♂♂, 2♀♀ sobre Q. rotundifolia, 22.VI.2012, 1♂, 2♀♀ sobre Q. rotundifolia, Ubrique, 25.V.2012, 1♀ sobre Q. suber. Cáceres, Monfragüe, 410 m, 3.VI.1997, 1 ej., A.Gruppe (AG). Cuenca, Uña, 40°13’15.71’’N, 1°58’50.71’’O, 1142 m, 27.VI.2009, 1♂ sobre Buxus sempervirens. Granada, La Herradura, Cantarriján, 30SVF36, 10 m, 9.VII.2013, 1♂ sobre P. lentiscus, 30.VII.2013, 2♂♂ sobre P. lentiscus, La Herradura, Cerro Gordo, 27.VII.2013, 2♂♂ sobre P. lentiscus, La Herradura, Punta de la Mona, 100 m, 24.IV.2011, 1♂ sobre Acacia dealbata, 5.VII.2015, 1♂ sobre P. lentiscus, 2♂♂ sobre O. europaea, 7.II.2015, 1♂ a la luz, Cáñar, 30SVF6289, 1580 m, 18.VII.2011, 1♂, 1♀ sobre Q. pyrenaica. Huelva, Castaño de Robledo, 23.V.2012, 1♂ sobre Q. canariensis, Matalascañas, 20.VI.2012, 1♂ sobre Acacia sp., Patrás, 19.VI.2012, 1♂, 1♀ sobre P. lentiscus, Puerto Blanco, 19.VI.2012, 3♂♂, 5♀♀ sobre Eucaliptus globulus. Madrid, Chinchón, El Salitral, 40°10’73’’N, 3°28′76’ O, 560 m, 20.VI.2009, 1♀ a la luz, Ciudad Universitaria, 26.IV.2007, 1♀ volando
entre arbustos, 10.VI.2008, 1♀ sobre Q. robur, 9.V.2012, 1♀ volando al amanecer, 9.VI.2014, 5♂♂, 5♀♀ sobre Q. robur. Málaga, Algarro, Finca La Mayora, 30S 407240, 92 m, 8.V.2007, 1♂, 5.VII.2007, 1♀, 12.IX.2007, 1 larva, 13.XI.2007, 1♂, 4♀♀, 3 larvas,
todos recolectados con paraguas japonés vareando aguacates, José Miguel Vela (IFAPA), 3.VI.2008, 1♂, 17.VI.2008, 1♀, 17.VII.2008,
1♂, 6.VIII.2008, 1♂, 19.VIII.2008, 1♀, todos en trampas de intersección y de caída colgadas en aguacates, José Miguel Vela,
Coín, Sa. Alpujata, 3.V.2013, 5♂♂, 4♀♀ sobre Salix pedicellata, Cómpeta, 7.IV.2007, 1♂, 1♀ sobre C. siliqua, El Madroñal, Embalse del Río Guadaiza, 13.VI.2013, 1♂, 1♀ sobre S. pedicellata, Estepona, Sa. Bermeja, Río Padrón, 12.VI.2013, 7♂♂, 12♀♀ sobre S. pedicellata, Vélez Málaga, Finca La Alegría, 30S 397065, 140 m, 19.IX.2007, 4♀♀, 2 larvas, 25.IX.2007, 1 larva, 4.X.2007, 2♀♀, 7 larvas,
17.X.2007, 2♀♀, 19.XI.2007, 1♂, 2 larvas, 27.XI.2007, 1♀, 13.XII.2007, 1♀, 1 larva, 20.XII.2007, 1♂, 1♀, 1 larva, 10.I.2008,
1♀, 4.II.2008, 1♂, 1♀, 26.II.2008, 2♂♂, 2♀♀, 3.IV.2008, 1♀, todos recolectados con paraguas japonés vareando aguacates, José
Miguel Vela (IFAPA), 19.V.2008, 1♂, 2♀♀, 17.VI.2008, 1♀, 4.VII.2008, 1♂, 17.VII.2008, 1♀, 8.I.2009, 2♂♂, 25.I.2009, 1♂, 27.II.2009,
3♂♂, 1♀, 11.III.2009, 1♂, 2♀♀, 24.III.2009, 1♂, 14.IV.2009, 1♂, 11.V.2009, 1♀, todos en trampas de intersección y de caída
colgadas en aguacates, José Miguel Vela, Llanos de Purla, 2.V.2013, 2♀♀ sobre S. pedicellata. Murcia, Jumilla, Sa. del Carche, 990 m, 22.VII.2008, 1 ej., Lecina & Gallego (AG), Miranda, 28.VIII.2005, 1♂, 1♀ sobre C. siliqua. Pontevedra, Fontán, 42°30’37.30’’N, 8°31’21.39’’O, 194 m, 21.IX.2009, 4♂♂, 9♀♀ sobre Q. robur, Pereiro, 42°26’40.89’’N, 8°42’32.16’’O, 150 m, 20.IX.2009, 1♂ sobre Q. robur. Tarragona, La Selva del Camp, 41°13′07.67′′N, 1°08′36.94′′E, 224 m (por su elevado número, solo anotamos parte del material estudiado)
20.XI.2005, 2♂, 4♀♀, 7 larvas, 27.XII.2005, 1♀, 2 larvas, 20.I.2007, 1 larva, 19.II.2007, 1♂, 2♀♀, 2 larvas, 18.III.2007,
1♂, 15.IV.2007, 1♀, 1 larva, 18.VI.2007, 1♂, 14.VII.2007, 1 larva, 13.VIII.2007, 1 larva, 15.IX.2007, 1♀, 13.X.2007, 1♂, 8♀♀,
14.XII.2007, 1 larva, 12.II.2008, 4♂♂, 2♀♀, 22.II.2008, 1♂, 8.III.2008, 1♂, 1♀, 7.IV.2008, 1♂, 1♀, 14.VII.2008, 1♀, 1 larva,
21.VIII.2008, 2♂♂, 1♀, 15.IX.2008, 3♂♂, 3♀♀, 1 larva, 21.X.2008, 5♀♀, 1 larva, 18.XI.2008, 8 larvas, 12.XII.2008, 1♀, 1 larva,
27.I.2010, 2♂♂, 22.II.2010, 2♂♂, 2♀♀, 1 larva, 20.III.2010, 4♂♂, 23.III.2010, 1♂, 23.IV.2010, 1♀, 17.VIII.2010, 2♂♂, 8♀♀,
4 larvas, 22.IX.2010, 1♂, 4♀♀, 9 larvas, 20.X.2010, 1♂, 5♀♀, 3 larvas, 19.XI.2010, 4♀♀, 1 larva, 14.XII.2010, 1♂, 3♀♀, 1 larva,
15.I.2011, 1♀, 4 larvas, 13.II.2011, 1♀, 1 larva, 10.IV.2011, 1♂, 14.IV.2011, 1♀, 14.V.2011, 1♀, 16.VII.2011, 2♀♀, 1 larva
12.VIII.2011, 1♀, 6.IX.2011, 2♂♂, 4♀♀, 2 larvas, 15.X.2011, 3♂♂, 10♀♀, 12.XI.2011, 2♀♀, 2 larvas, 16.XII.2011, 1♂, todos sobre
Citrus clementina, N. Cañellas. ITALIA: Sardegna, Nuoro, Fonni, 40°07′09′′N, 9°15′16′′E, 971 m, 21.V.2008, 1♂ sobre Castanea sativa. PORTUGAL: Alto Alentejo, Évora, 240 m, 24.VI.2000, 1 ej., A. Gruppe (AG). Beira Interior, Guarda, Sra. de Pestero, 800 m, 20.VI.2011, 27 ej., A. Gruppe (AG). Beira Litoral, Aveiro, Buçaco, 440 m, 20.VI.2011, 6 ej., A. Gruppe (AG). Braga, PN Geres, Vilar de Veigna, 17.VI.2011, 1 ej., A. Gruppe (AG). Minho, Viana do Castelo, PN Serra da Penada, Lamas de Mouro, 17.VI.2011, 1 ej., A. Gruppe (AG). Trás-os-Montes e Alto Douro, Vila Real, Sa. Do Alvão, Lomas de Olo, 1000 m, 18.VI.2011, 1 ej., A. Gruppe (AG).
Hemisemidalis pallida (Withycombe, 1924) (Figs. 37, 43, 53-60, 267-274)
Parasemidalis pallida Withycombe, 1924
Withycombe, 1924a: 141 (d).
Hemisemidalis pallida (Withycombe, 1924)
Monserrat, 1976b: 247 (rf), 249 (dis), 1977: 185 (nt); Aspöck et al., 1980: 158 (bio, mf, tx, rf); Monserrat, 1983b: 126 (nt), 1985b: 91 (bio, dis), 1985d: 135 (bio, dis); Díaz-Aranda et al., 1986b: 1143 (bio, dis); Marín & Monserrat, 1987: 350-359 (bio, dis); Monserrat, 1987a: 145 (bio, dis); Monserrat & Díaz-Aranda, 1987: 185 (bio, dis); Díaz-Aranda & Monserrat, 1988a: 121 (bio, dis), 1988b: 227 (bio, dis), 1988c: 224 (bio, dis); Monserrat & Díaz-Aranda, 1990: 46 (bio, dis); Marín & Monserrat, 1990: 228 (bio, dis), 1991b: 183 (bio, dis); Monserrat & Marín, 1992: 283 (bio); Marín, 1994: 260-297 (bio, dis, fen); Monserrat et al., 1994: 59 (bio, dis); Marín & Monserrat, 1995a: 45 (bio, dis), 1995b: 119 (bio, dis); Monserrat, 1995: 123 (bio, dis), 1996a: 15 (list); Lo Verde & Monserrat, 1997: 61 (rf); Dessart, 1998: 404 (bio, dis); Monserrat et al., 1999: 40, 42 (bio, dis); H. Aspöck et al., 2001: 192 (rf); Monserrat, 2002: 13 (bio, dis), 2005a: 27 (bio, dis, tx); Sziráki & Harten, 2006: 430 (rf); Monserrat, 2010: 27 (bio, dis, tx), 2013: 303 (bio, dis); Monserrat et al., 2013: 54 (bio, dis); Monserrat & Triviño, 2013: 20 (list), 130 (dis).
Especie conocida del límite meridional de la Región Paleártica occidental, desde Mongolia, Afganistán, Usbekistán, Kazakhstán,
Pakistán, Jammu, Cachemira, Irak, Irán, Yemen, Arabia Saudí, Líbano, Anatolia, etc., a Egipto, Sudán, Argelia y Marruecos
en el norte de África, y España, Italia y Grecia en Europa. Fuertemente asociada a estepas, zonas térmicas y secas, y citada
sobre plantas de medios subdesérticos y fanerófitos xerófilos en medios de influencia mediterránea (Tamarix, Haloxylon, Nerium, Pistacea, Tetraclinis, Caragana, Olea, Acacia), también sobre Pinus, Quercus, Cupressus y Eucaliptus, y herbáceas (Lasiagrostis y Ephedra), pero sobre todo asociada a Juniperus spp. Recolectada prácticamente durante todo el año, según zonas, en altitudes de hasta 2900 m. En nuestra fauna presenta
una tendencia extra-eurosiberiana, ha sido mayoritariamente recolectada a la luz (puede llegar a ser puntualmente muy abundante),
y ha sido citada especialmente sobre enebros o sabinas, sobre los que se han recolectado la mayoría de los ejemplares con
datos sobre plantas, y con ejemplares emergidos de frutos, gálbulos/conos o agallas de Dendrocerus indicus (Hymenoptera: Megaspilidae) y Oligotrophus sp. (Diptera: Cecidomiidae) en enebros y sabinas, también sobre Pinus, Salix, Quercus, Tamarix, vegetación herbácea y en espacios urbanos, entre IV-X (Marín, 1994 y Monserrat, 1995, a partir de muestreos seriados y de estudios de seguimiento con trampas parecen demostrar para esta especie tener un ciclo
monovoltino), en altitudes que oscilan entre 10-1900 m.
La genitalia masculina de esta especie está sometida a una cierta variabilidad, especialmente en la extensión y límite del
margen caudal del hipandrio (Monserrat, 2010). En la figuras 267-270 anotamos los extremos encontrados en la variabilidad hallada entre los diversos ejemplares ibéricos
estudiados, siendo la que se anota en la fig. 269 la que poseen la mayoría de los ejemplares. Sin tener en cuenta esta variabilidad,
este carácter ha sido utilizado con frecuencia para describir numerosas especies paleárticas que, sin duda, pertenecen a la
especie que tratamos (Monserrat, 2010).
Los estadios preimaginales (Figs. 37, 43, 53-60) fueron descritos por Monserrat & Hölzel (1987: 136).
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Alicante, Olmeda del Prado, 560 m., 5.VIII.2008, 1 ej., Lencina (AG). Almería, San José, Playa de Mónsul, 28.VI.2011, 1♀ a la luz, 20.VII.2011, 1♂, 1♀ a la luz, Tabernas, Rambla Roja, 30SWG50, 400 m,
7.VII.2013, 5♂♂, 3♀♀ sobre Tamarix sp., F. Acevedo, 6♂♂, 1♀ a la luz. Cuenca, Beteta, 40°34’37.40’’N, 2°02’33.10’’O, 1310 m, 27.VI.2009, 12♂♂, 1♀ volando al atardecer sobre Juniperus nana, Verdelpino de Cuenca, El Chantre, 40°07’42.56’’N, 2°06’53.88’’O, 948 m, 27.VI.2009, 1♀ sobre J. sabina, Villalba de la Sierra, 40°14’12.42’’N, 2°05’23.30’’O, 1000 m, 27.VI.2009, 1♂ sobre J. oxycedrus, 1♂, 4♀♀ sobre J. sabina. Granada, Punta de la Mona, 30SVF36, 200 m, 10.VIII.2010, 2♂♂ a la luz. Huelva, Matalascañas, 24.V.2012, 3♂♂, 2♀♀ sobre J. phoenicea.
Nimboa adelae Monserrat, 1985 (Figs. 243-249, 305)
Monserrat, 1985d: 137 (d), 1987a: 144 (bio, dis), 145 (nt); Monserrat & Díaz-Aranda, 1990: 41 (dis); Meinander, 1990: 36 (rf); Monserrat & Marín, 1992: 282 (bio); H. Aspöck et al., 2001: 176 (rf); Sziráki, 2004: 21 (rf); Monserrat, 2013: 303 (rf) ; Monserrat & Triviño, 2013: 20 (list), 130 (dis).
Especie anteriormente conocida del S. E. de España (Murcia y Almería), ahora la citamos del sur de Granada, y ha sido hallada
(Monserrat & El-Serwy, 2010) y ahora citamos en Egipto depredando sobre colonias de Pulvinaria tenuivalvata (Newstead) (Hemiptera: Coccidae), por lo que una distribución mucho más amplia por la región mediterránea meridional cabe
presumirse. Está mayoritariamente citada entre X-XII, en altitudes de 10-30 m. En nuestra fauna ha sido citada mayoritariamente
a la luz en medios extremadamente xéricos y térmicos, carentes de vegetación arbórea, con cañizares o nanofanerófitos leñosos
y halófilos, entre VI-IX (probablemente sufra diapausa estival), en altitudes que oscilan entre 20-670 m.
Que sepamos, los estadios preimaginales son desconocidos.
NUEVO MATERIAL Y MATERIAL COMPLEMENTARIO ESTUDIADO: EGIPTO: Middle Egypt, Giza Governorate, Al-Ayyat, 40 km South Cairo, X-XII.2008, 1♂, 9♀♀, S. A. El-Serwy (VM), XII.2008, 2♂♂, S. A. El-Serwy (VM), Atfieh, XI.2006,
3♂♂, 1♀, S.A. El-Serwy (VM), todos sobre colonias de Pulvinaria tenuivalvata (Newstead) infestando caña de azúcar Saccarum officinarum L. ESPAÑA: Granada, La Herradura, Punta de la Mona, 100 m, 4.VII.2015, 1♂ sobre Olea europea, Lobres, 23.VII.2016, 1♀ sobre Arundo donax.
Nimboa espanoli Ohm, 1973 (Figs. 250-255)
Ohm, 1973a: 241 (d); Meinander, 1975b: 248 (rf), 1977: 84 (rf); Monserrat, 1978a: 67 (bio, rf); Aspöck et al., 1980: 9 (dis), 157 (bio, mf, tx, rf); Monserrat, 1983b: 126 (rf), 1985b: 90 (bio, dis), 1985d: 139 (nt), 1985e: 76 (rf), 1987a: 143 (bio, dis); Marín & Monserrat, 1987: 350-359 (bio, dis); Díaz-Aranda & Monserrat, 1988c: 224 (bio, dis); Meinander, 1990: 37 (rf); Monserrat & Marín, 1992: 282 (bio); Monserrat, 1995: 114 (dis); H. Aspöck et al., 2001: 176 (rf); Monserrat, 2002: 8 (bio, dis), 2010: 28 (rf), 2013: 303 (rf); Sziráki & Harten, 2006: 394 (rf); Monserrat & Triviño, 2013: 20 (list), 131 (dis).
Nimboa espanioli Ohm, 1973 (lapsus)
Sziráki, 2004: 22 (rf); Monserrat & Triviño, 2013: 20 (list).
Nimboa sp.
Meinander, 1972a: 189 (rf); Monserrat & Triviño, 2013: 20 (list).
A esta especie pertenece la cita de Meinander (1972a) y también deben pertenecer mayoritariamente las citas dadas en la zona mediterránea como Nimboa sp., que se han mencionado con anterioridad en la bibliografía.
Especie conocida del S. E. de España, Isla de Rodas, Islas Canarias, Marruecos, Mali, Nigeria, Sudáfrica (Transvaal) y Yemen.
Escasamente conocida, ha sido mayoritariamente recolectada a la luz en zonas áridas y térmicas, en medios xéricos y carentes
de vegetación arbórea, también sobre fanerófitos o nanofanerófitos de zonas xéricas, subdesérticas y mediterráneas (Chamarops humilis, Pinus halepensis, Juniperus oxycedrus, Cupressus sempervirens, Quercus rotundifolia, Ceratonia siliqua,
Pistacea lentiscus, P. terebinthus, Echinospartum boissieri y Nerium oleander). En África subsahariana ha sido citada sobre algodón, Baissea multiflora, Combretum micranthum y Dicrostochys glomerata. Está mayoritariamente citada entre II, VI-XI, en altitudes de 350-1.800 m. En nuestra fauna ha sido citada sobre Chamaerops, Nerium, Echinospartum, Quercus, Ceratonia, Pistacea, Pinus, Juniperus, Cupressus, entre V-IX, en altitudes que oscilan entre 10-1200 m.
Que sepamos, los estadios preimaginales son desconocidos.
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Almería, El Molino de La Serena, Bédar, 17.VII.2007, 1♂ a la luz, San José, 25.V.2003, 2♂♂, 1♀ a la luz. Málaga, Navahermosa, 26.V.2012, 1♀ sobre Olea europea.
Parasemidalis fuscipennis (Reuter, 1894) (Figs. 256-259)
Coniopteryx fuscipennis Reuter, 1894
Reuter, 1894: 13 (d).
Parasemidalis fuscipennis (Reuter, 1894)
Meinander, 1972a: 283 (rf); Aspöck et al., 1980: 156 (bio, mf, tx, rf); Ohm, 1986: 330 (rf); Popov, 1986: 169 (rf); Díaz-Aranda et al., 1986b: 1143 (bio, dis); Monserrat & Díaz-Aranda, 1987: 185 (bio, dis), 1990: 43 (bio, dis), 45, 46 (rf); Monserrat & Marín, 1992: 283 (bio); Monserrat, 1995: 123 (dis), 1996a: 15 (list); H. Aspöck et al., 2001: 190 (rf); Monserrat, 2002: 12 (bio, dis); Canbulat, 2007: 304 (rf); Monserrat & Triviño, 2013: 20 (list), 131 (dis).
Especie conocida de Europa, extendida a Anatolia, Ucrania y Mongolia. También citada de USA (Michigan, Arizona, California)
y México (Durango). Generalmente asociada a coníferas (Pinus y Juniperus), aunque también sobre planifolios (Quercus robur, Q. rotundifolia, Q. faginea, Carpinus, Olea europaea, Crataegus monogyna y Krascheninnikovia ceratoides), vegetación de rivera (Typha) y árboles en parques, entre IV-VIII, en altitudes de hasta 1520 m. En nuestra fauna tiende a ocupar zonas térmicas y secas,
y también ha sido escasamente citada sobre robles, olivos y endrinos, ocasionalmente sobre Krascheninninkovia, entre V-VIII, Monserrat (2002) cita algún ejemplar (imago) en I, lo que sugiere que pueda presentar actividad en invierno, en altitudes que oscilan entre
10-1350 m.
Como Parasemidalis annae Withycombe (1922a: 171; 1923: 588) describe el huevo.
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Granada, La Herradura, Cerro Gordo, 6.VIII.2011, 1♀ a la luz.
Parasemidalis triton Meinander, 1976 (Figs. 260-261)
Meinander, 1976: 87 (d); Monserrat & Díaz-Aranda, 1990: 44 (bio, dis, tx); Monserrat & Marín, 1992: 283 (bio); Monserrat, 1995: 123 (bio, dis); H. Aspöck et al., 2001: 191 (rf); Sziráki, 2004: 39 (rf); Monserrat & Triviño, 2013: 20 (list), 132 (dis).
La validez de esta especie frente a Parasemidalis fuscipennis (Reuter, 1894) ha sido puesta en duda en alguna ocasión (Monserrat & Díaz-Aranda, 1990), dada la variabilidad encontrada en algunos caracteres genitales, especialmente en la forma caudal del hipandrio masculino,
hecho también ocurre en las especies del género Hemisemidalis (Figs. 268-270).
Elemento muy escasamente conocido, descrito de Argelia a partir de un único ejemplar, puntualmente citado en la Península
Ibérica (Guadalajara). Está citada en II, a 212 m de altitud. Los escasos ejemplares hallados en nuestra fauna parecen ajustarse
a la descripción original de esta especie (Meinander, 1976). En ella ha sido citada sobre Quercus faginea y Juniperus thurifera, entre V-VI, en altitudes que oscilan entre 1179-1190 m.
Que sepamos, los estadios preimaginales son desconocidos.
Semidalis aleyrodiformis (Stephens, 1836) (Figs. 283-285, 310)
Coniopteryx aleurodiformis Stephens, 1836
Stephens, 1836: 116 (d); McLachlan, 1868: 193 (dis), 1880: 106 (dis); Löw, 1885: 84 (rf); Dziędzielewicz, 1905: 383 (rf); Monserrat & Triviño, 2013: 20 (list).
Semidalis aleyrodiformis (Stephens, 1836)
Klapálek, 1909: 30 (dis); Killington, 1936: 211 (rf); Eglin, 1940: 270 (rf); Meinander, 1972a: 318 (rf); Monserrat, 1977a: 23 (list), 181 (key, mf), 183 (bio, dis), 1978a: 65 (bio, dis), 1979a: 415 (bio, dis), 1978b: 182 (bio, dis); Aspöck et al., 1980: 159 (bio, mf, tx, rf); Monserrat, 1980: 192 (bio, dis), 1982a: 83 (bio, dis), 1984a: 148 (dis, tx), 1984b: 108 (bio, dis), 1984c: 160 (bio, dis), 1984d: 44 (bio,
dis), 1984e: 176 (bio, dis), 1984g: 32 (bio, dis), 1985a: 96 (bio, dis), 1985b: 90 (bio, dis), 1985d: 135 (bio, dis); Díaz-Aranda et al., 1986a: 1125 (bio, dis), 1986b: 1143 (bio, dis); Monserrat, 1987a: 144 (bio, dis); Marín & Monserrat, 1987: 350-359 (bio, dis); Monserrat & Díaz-Aranda, 1987: 184 (bio, dis), 1988a: 93 (bio, dis); Díaz-Aranda & Monserrat, 1988a: 121 (bio, dis), 1988b: 227 (bio, dis), 1988c: 225 (bio, dis); Monserrat & Díaz-Aranda, 1989: 55 (bio, dis); Marín & Monserrat, 1989: 296 (bio, dis); Monserrat & Díaz-Aranda, 1990: 46 (bio, dis); Marín & Monserrat, 1991b: 183 (bio, dis); Monserrat & Marín, 1992: 283 (bio); Marín, 1994: 260-297 (bio, dis, fen); Monserrat et al., 1994: 59 (bio, dis); Marín & Monserrat, 1995a: 45 (bio, dis), 1995b: 120 (bio, dis); Monserrat, 1995: 124 (bio, dis), 1996a: 15 (list); Monserrat et al., 1999: 42 (bio, dis); H. Aspöck et al., 2001: 195 (rf); Monserrat, 2002: 14 (bio, dis); Alvis et al., 2003: 186 (bio, dis); Monserrat, 2005a: 74 (rf); Zimmermann, 2005b: 37 (dis); Letardi, 2012: 72 (dis); Ocharan et al., 2012: 246 (rf); Monserrat, 2013: 303 (bio, dis), 305 (rf); Monserrat et al., 2013: 54 (bio, dis); Monserrat & Triviño, 2013: 20 (list), 132 (dis); Letardi et al., 2013: 36 (list).
Semidalis aleurodiformis (Stephens, 1836)
Enderlein, 1906: 212 (rf); Navás, 1908: 129 (dis, mf); Macho Bariego, 1909: 75 (dis); Navás, 1910a: 41, 45, 46 (dis), 1911: 209 (dis), 1913a: 85 (dis), 1913b: 68 (dis), 1914c: 41 (dis), 1914d: 190 (dis), 1915: 30 (dis),
1916a: 175 (dis); Bohigas & Sanchez, 1917: 318 (dis); Navás, 1919a: 202 (dis), 1921b: 65 (dis), 1923b: 8 (dis), 1924a: 246 (dis, key, mf), 1925a: 124 (dis, key, mf), 1925b: 138 (dis), 1927b: 98 (dis); Stitz, 1927: 18 (rf); Navás, 1928: 102 (dis), 1929a: 33 (dis); Esben-Petersen, 1931: 5 (dis); Berland, 1962: 27 (rf); Monserrat, 2013: 305 (rf); Monserrat & Triviño, 2013: 20 (list).
Semidalis curtisiana Enderlein, 1906
Enderlein, 1906: 212 (d); Navás, 1908: 33 (rf); Klapálek, 1909: 31 (rf); Navás, 1910a: 40, 41, 45, 54 (dis), 1914c: 41 (dis), 1914d: 190 (dis), 1915: 52 (dis), 1916a: 175 (dis), 1916b: 155 (dis), 1916c: 190
(dis); Bohigas & Sanchez, 1917: 318 (dis); Navás, 1919a: 202 (dis), 1919b: 43 (dis), 1921a: 152 (dis), 1921b: 65 (dis), 1924a: 246, 247 (dis, key, mf), 1925a: 124, 125 (dis, key, mf), 1927b: 98 (dis), 1928: 102 (dis), 1931: 121 (dis); Monserrat, 1977a: 23 (list), 185 (nt); Monserrat & Triviño, 2013: 20 (list).
Especie Paleártica expansiva, extremadamente eurioica, generalmente asociada con medios arbóreos, tanto planifolios como
aciculifolios, citada sobre Abies, Pinus, Juniperus, Cupressus, Populus, Corylus, Fagus, Quercus, Celtis, Crataegus, Carpinus, Ulmus, Prunus, Ceratonia,
Citrus, Acacia, Ilex, Pistacea, Buxus, Arbutus, Castanea, Olea, Taxus, Pyrus, Retama, Arundo, Zizyphus, etc. Es una de las especies europeas de coniopterígidos más eurioica, viviendo desde bosques húmedos subalpinos a zonas
subdesérticas, entre IV-X, en altitudes de hasta 2000 m. La extrema variabilidad morfológica en una especie con tan amplia
distribución (Meinander, 1972a) induce a pensar que probablemente tratamos un grupo de especies. En nuestra fauna ha sido ampliamente citada, incluido Baleares
(Mallorca), sobre una infinidad de sustratos vegetales, sean coníferas, o especialmente planifolios, tanto de naturaleza eurosiberiana
como continental o mediterránea, también en espacios urbanos y cultivos, mayoritariamente entre II-XI, en altitudes que oscilan
entre 10-1890 m. Monserrat (2002) anotaba actividad, tanto de imagos como de larvas en XI, XII sobre cítricos en Valencia, y los datos ahora anotados en el
nuevo material recolectado en estas zonas de influencia mediterránea permiten sugerir actividad durante todo el año, incluso
durante el invierno, tanto de larvas, como de imagos, al menos en las zonas mediterráneas.
Los estadios preimaginales de esta especie fueron descritos o reiterados por Withycombe (1923: 585), Lacroix (1924: 79), Withycombe (1925a: 410; 1925b: 20), Stitz (1931: 101), Killington (1936: 208), Silvestri (1942: 30, 31), Collyer (1951: 563), Yang (1951: 342), Fraser (1959: 33), Hoffmann (1962: 327), Rousset (1966: 34, 1969: 110), Lauterbach (1972: 143), Agekyan (1978: 509), Gepp & Stürzer (1986: 241-257), Starý et al. (1988: 66); Gepp (1999: 194, 199). Sobre los estadios preimaginales de esta especie podemos añadir que una de las hembras ahora citada (colectada
en Madrid, Ciudad Universitaria, 29.V.2009) efectuó una puesta de 11 huevos translúcidos, todos sobre el margen de las hojas que fueron
incluidas en el recipiente donde fue mantenida, bien aislados o agrupados de dos en dos (Figs. 3, 4). Su desarrollo empleó 6 días, a 23-39 °C, desde el día de su puesta a la emergencia de las larvas. Su micropilo era de color
blanco intenso, y el corion portaba depresiones poligonales de las células foliculares, como ha sido habitualmente citado
(Figs. 25a, b).
NUEVO MATERIAL Y MATERIAL COMPLEMENTARIO ESTUDIADO: ESPAÑA: Álava, Iruraitz Gauna, Gazeo, 30TWN4644, 600 m, 25.V.2007, 1♀ sobre Crataegus monogyna, S. Pagola. Almería, Cueva de Sorbes, 1.V.2007, 1♂ sobre Olea europaea, El Molino de La Serena, Bédar, 1.V.2007, 1♀ sobre Pistacea sp., 7♂♂, 3♀♀ sobre Citrus sp., 8♂♂, 13♀♀ sobre Rhamnus alaternus, 14.VII.2007, 2♂♂, 1♀ sobre Ficus carica, La Rambla Honda, 30SWF7497, 597 m, 7.V.2011, 1♂ sobre O. europea, La Serena, Bédar, 2.V.2007, 3♂♂, 2♀♀ sobre Ceratonia siliqua, 13.VII.2007, 1♂ sobre R. lycioides, 14.VII.2007, 2♂♂ sobre C. siliqua, 17.VII.2007, 3♂♂, 1♀ sobre O. europaea, 3♂♂, 4♀♀ sobre Pistacea lentiscus. Ávila, Navarredonda de Gredos, 28.VII.2007, 1♂ sobre Quercus pyrenaica. Badajoz, Mérida, 29.IV.2006, 6♂♂, 5♂♂ volando al amanecer sobre Rosa sp., 29.IV.2006, 3♂♂ sobre Citrus limon. Cádiz, Barbate, Caños de Meca, 20.VI.2012, 1♂ sobre O. europaea, Bolonia, 21.VI.2012, 1♂, 2♀♀ sobre P. lentiscus, 3♂♂ sobre O. europaea, Estación de la Almoraima, 25.V.2012, 1♂, 2♀♀ sobre Q. canariensis, 21.VI.2012, 1♂, 2♀♀ sobre Q. canariensis, Facinas, 20.VI.2012, 2♂♂, 3♀♀ a la luz, Grazalema, 31.V.2005, 770 m, 3 ej., A. Gruppe (AG). Parque Natural del Estrecho,
21.VI.2012, 2♂♂ sobre O. europaea, Pinsapar de Grazalema, 3.V.2005, 1♂ sobre Q. canariensis, 1♂ sobre Arbutus unedo, 1♂ sobre Q. suber, 3♂♂ sobre Q. rotundifolia, 26.V.2012, 1♀ sobre Q. canariensis, 1♂, 1♀ sobre Abies pinsapo, Ubrique, 25.V.2012, 1♀ sobre Q. suber, 1♂♂, 8♀♀ sobre Q. canariensis. Cuenca, Puerto Cabrejas, 40°03’16.98’’N 2°20’45.76’’O, 1150 m, 27.VI.2009, 3♀♀ sobre Q. faginea. Granada, Aldeire, 30SVG9410, 1661 m, 8.V.2011, 1♂, 2♀♀ sobre Pinus pinaster, 1♀ sobre P. halepensis, Calahonda, 30SVF66, 20 m, 8.VII.2013, 4♀♀ sobre P. halepensis, Cáñar, 30SVF6289, 1580 m, 18.VII.2011, 1♂ sobre Q. pyrenaica, F. Acevedo, El Rescate, 3.VIII.2010, 1♂, 1♀ sobre O. europea, 23.VIII. 2010, 1♂, 1♀ sobre O. europea, 8.VIII.2011, 1♂, 1♀ sobre O. europea, 8.VIII.2011, 1♂, 1♀ sobre O. europea, Ferreira, 30SVG9513, 1356 m, 8.V.2011, 3♂♂, 1♀ sobre P. halepensis, Jete, 3.VIII.2010, 3♂♂, 1♀ sobre O. europea, 23.VIII. 2010, 2♀♀ sobre O. europea, 8.VIII.2011, 1♂, 1♀ sobre O. europea, 8.VIII.2011, 2♀♀ sobre O. europea, La Herradura, Cantarriján, 30SVF36, 10 m, 9.VII.2013, 1♀ sobre P. lentiscus, 1♀ sobre P. halepensis, 30.VII.2013, 1♂, 2♀♀ sobre P. lentiscus, La Herradura, Cerro Gordo, 1.VIII.2010, 1♀ sobre P. lentiscus, 7.VIII.2010, 2♂♂, 1♀ sobre P. lentiscus, 12-22.VIII.2010, 1♀ en trampa de Malaise, 24.IX.2010, 1♂ sobre P. lentiscus, 21.IV.2011, 1♂ sobre P. lentiscus, 6.VIII.2011, 2♀♀ a la luz, 13.VIII.2013 1♀ a la luz, 30.VIII.2013, 1♂ sobre P. lentiscus, La Herradura, Punta de la Mona, 30SVF36, 100 m, 26.VIII.2010, 1♀ a la luz, 4.VII.2015, 3♂♂ sobre O. europea, 5.VII.2015, 1♂ sobre C.siliqua, 25.IX.2015, 1♂, Gualchos, 30SVF6462, 30 m, 28.VI.2011, 1♀ sobre Melia azedarach, Lobras, 30SVF8084, 822 m, 6.V.2011, 1♀ sobre Q. rotundifolia, Los Tablones, 13.IV.2014, 6♂♂, 2♀♀ sobre O. europaea, Majadillas, 3.VIII.2010, 1♀ sobre Pistacea sp., 23.VIII. 2010, 1♂, 1♀ sobre N. holeander, 8.VIII.2011, 1♀ sobre Pistacea sp., 21.VIII. 2011, 1♂, 1♀ sobre N. holeander, Otivar, 3.VIII.2010, 3♂♂ sobre O. europea, 23.VIII. 2010, 1♂, 1♀ sobre O. europea, 8.VIII.2011, 3♂♂ sobre O. europea, 21.VIII. 2011, 1♂, 1♀ sobre O. europea, San Antonio, 7.IV.2007, 1♂, 2♀♀ a la luz, 3♀♀ sobre O. europaea, Vélez de Benaudalla, 350 m, 13.IV.2014, 1♂, 2♀♀ sobre O. europaea. Huelva, Castaño de Robledal, 23.V.2012, 1♂ sobre Castanea sativa, La Corte, 23.V.2012, 1♂ sobre P. lentiscus. Madrid, Ciudad Universitaria, 7.V.2007, 1♂ volando al amanecer, 27.V.2008, 1♂ volando al amanecer, 3.V.2009, 1♂ volando al amanecer,
29.V.2009, 1♀ volando al amanecer, 5.V.2010, 2♂♂, volando al amanecer, 19.V.2010, 3♂♂, 2♀♀ volando al amanecer, 26.IV.2011,
1♂, volando al amanecer, 12.V.2011, 1♂, volando al amanecer, 27.IV.2012, 6♂♂, 1♀ volando al amanecer, 9.V.2012, 7♂♂, volando
al amanecer, 22.V.2012, 7♂♂, volando al amanecer, 22.IV.2014, 1♂ volando al amanecer, 28.IV.2014, 4♂♂, volando al amanecer,
29.IV.2014, 6♂♂, volando al amanecer, 5.V.2014, 4♂♂, volando al amanecer, El Escorial, Abantos, Fuente de la Teja, 21.V.2008,
1♀ sobre A. pinsapo, Fuente del Cervunal, 1.700 m, 24.VII.2007, 1♂ sobre P. sylvestris. Málaga, Algarrobo, 5.VIII.2010, 1♂, 3♀♀ sobre C. siliqua, 25.VIII.2010, 1♂ sobre O. europea, Benamocarra, 5.VIII.2010, 2♂♂, 3♀♀ sobre O. europea, 25.VIII.2010, 1♂ sobre O. europea, 25.VIII.2010, 1♂ sobre P. halepensis, 10.VIII.2011, 1♂, 3♀♀ sobre C. siliqua, 30.VIII.2011, 1♂ sobre O. europea, Benamocarra, 10.VIII.2011, 2♂♂, 3♀♀ sobre O. europea, 30.VIII.2011, 1♂ sobre O. europea 30.VIII.2011, 1♂ sobre P. halepensis, Canillas de Albaida, Fábrica de la luz, Río de la Llanada de Turvilla, 30S 413487 4080230, 720 m, 28.VI.2015, 1♀, C. Zamora, Coín, Sa. Alpujata, 3.V.2013, 5♂♂, 4♀♀ sobre Salix pedicellata, 1.V.2014, 6♂♂, 3♀♀ sobre O. europea, Cómpeta, 7.IV.2007, 1♂ sobre C. siliqua, Frigiliana, 340 m, 2.V.2014, 6♂♂, 3♀♀ sobre O. europea, Las Asomadillas, 25.V.2012, 1♂, 2♀♀ sobre P. lentiscus, Llanos de Purla, 2.V.2013, 1♀ sobre S. pedicellata, Maro, 2.V.2014, 2♂♂, 3♀♀ sobre O. europea, 1♂, 3♀♀ sobre P. halepensis, Vélez Málaga, Finca La Alegría, 30S 397065, 140 m, 11.III.2009, 1♂, 11.V.2009, 1♂ en trampas de intersección y de caída colgadas
en aguacates, José Miguel Vela, Vélez Málaga, Sayalonga 5.VIII.2010, 5♂♂ sobre O. europea, 25.VIII.2010, 1♂ sobre O. europea, 10.VIII.2011, 5♂♂ sobre O. europea, 25.VIII.2010, 1♂ sobre O. europea. Murcia, Jumilla, Sa. del Carche, 990 m, 19.IX.2008, 1 ej., Lecina & Gallego (AG), Miranda, 28.VIII.2005 5♂♂, 5♀♀ sobre C. siliqua, Mijas, 1.V.2014, 2♂♂, 3♀♀ sobre O. europea, Pinilla, 10.VII.2005 1♂, 1♀ sobre P. halepensis. Segovia, Valsaín, 8.VII.2007, 1♀ sobre P. sylvestris. Tarragona, La Selva del Camp, 41°13′07.67′′N, 1°08′36.94′′E, 224 m (por su elevado número, solo anotamos parte del material estudiado),
27.XII.2005, 1♂, 1♀, 2 larvas, 15.IV.2007, 1♀, 14.VII.2007, 1♀, 7.IV.2008, 1♂, 1♀, 14.VII.2008, 1♀, 21.VIII.2008, 1♀, 17.VII.2010,
2♂♂, 20.X.2010, 1♀, 14.V.2011, 1♀, 16.VII.2011, 1♂, 3♀♀, 12.VIII.2011, 1♀, todos sobre C. clementina, N.Cañellas. Vizcaya, Bilbao, 25.IX.2011, 1♀ sobre Quercus sp., en parque, Plentzia, 23.IX.2011, 1♀ errante. Zaragoza, Daroca, 17.VI.2007, 1♀ sobre P. halepensis, Salvatierra, 23.VIII.2012, 1♀ sobre vegetación de rivera. ITALIA: Liguria, Génova, V.1900 1♀, R. Gestro (MG). Sardegna, Sassari, Baldinca, 23.V.2008, 3♂♂, 2♀♀ sobre O. europaea, Chighizzu, 19.V.2008, 1♂, 1♀ sobre R. alaternus, Prato Comunale, Rio Manno, 40°41′14′′N, 8°28′58′′E, 57 m, 18.V.2008, 1♀ sobre P. lentiscus. PORTUGAL: Alto Alentejo, Évora, 240 m, 24.VI.2000, 1 ej., A. Gruppe (AG). Beira Interior, Guarda, Sra. de Pestero, 800 m, 20.VI.2011, 3 ej., A. Gruppe (AG). Braga, PN Geres, Vilar de Veigna, 17.VI.2011, 1 ej., A. Gruppe (AG).
Semidalis pluriramosa (Karny, 1924) (Figs. 64-69, 275-278, 289-290)
Protosemidalis pluriramosa Karny, 1924
Karny, 1924: 476 (d).
Semidalis pluriramosa (Karny, 1924)
Monserrat & Díaz-Aranda, 1990: 47 (bio, dis); Monserrat et al., 1991: 109 (rf); Monserrat & Marín, 1992: 283 (bio); Monserrat, 1996b: 19 (rf); H. Aspöck et al., 2001: 195 (rf); Sziráki, 2004: 45 (rf); Monserrat, 2005a: 30 (bio, dis); Sziráki & Harten, 2006: 431 (rf); Monserrat, 2006: 20 (dis), 23 (dis), 2010: 28 (rf), 30 (bio, dis), 2013: 303 (rf); Monserrat & Triviño, 2013: 20 (list), 133 (dis).
Especie circunsahariana expansiva, poco citada, y conocida de Egipto, Argelia, Sudán, Nigeria, Marruecos, Túnez, Mali, Senegal,
Zimbabwe y Zambia, extendida desde las zonas más secas y térmicas y subdesérticas del S. E. de la Península Ibérica en el
oeste y hasta Yemen hacia el este. Mayoritariamente citada en medios xéricos sobre muy diversos sustratos vegetales (Eucalyptus, Citrus, Phoenix, Olea, Ceratonia, Amygdalus, Acacia, Dicrostochys, Zizyphus, Tamarix, Arundo, Vitellama, Combretum, Baissea, Brachystegia, Melia y Tetraclinis), también muy frecuentemente recolectada a la luz en zonas secas, ramblas secas (con Tamarix gallica y Arundo donax) y en zonas salinas con vegetación halófila (Chenopodiaceae), y ha sido recolectada prácticamente a lo largo de todo el año, según zonas, a veces a nivel del mar, hasta los 1700 m.
En nuestra fauna sólo ha sido puntualmente citada en el sudeste árido (sur de Almería y Granada) coincidiendo con los medios
citados, mayoritariamente recolectada a la luz, también sobre Pistacea, entre VII-VIII, en altitudes que oscilan entre 10-200 m.
Los estadios preimaginales de esta especie (Figs. 64-69) fueron descritos por Monserrat et al. (1991: 110).
NUEVO MATERIAL COMPLEMENTARIO ESTUDIADO: La genitalia del único ejemplar (♀) que citamos coincide con lo anotado para esta especie por Monserrat & Reviejo (1980). A la espera de poder recolectar ejemplares machos que confirmen esta identificación, sugerimos su posible presencia en
Cerdeña. ITALIA: Cerdeña, Sassari, Belvi, Scala Ceresia, 39°57′19′′N, 9°10′53′′E, 665 m, 21.V.2008, 1♀ sobre Erica arborea.
Semidalis pseudouncinata Meinander, 1963 (Figs. 70-75, 286-288, 309)
Meinander, 1963: 103 (d), 1972a: 325 (dis, mf); Ohm, 1973b: 302 (rf); Gepp, 1974b: 167 (rf); Monserrat, 1977a: 23 (list), 181 (key, mf), 182 (bio, dis), 1978a: 65 (bio, dis), 1979a: 415 (bio, dis); Aspöck et al., 1980: 161 (bio, mf, tx, rf); Monserrat, 1984a: 149 (dis, tx), 1984b: 109 (bio, dis), 1984d: 44 (bio, dis), 1984g: 33 (bio, dis), 1985b: 90 (bio, dis), 1985d: 137 (bio, dis); Díaz-Aranda et al., 1986a: 1125 (bio, dis), 1986b: 1144 (bio, dis); Monserrat, 1987a: 145 (bio, dis); Monserrat & Díaz-Aranda, 1987: 184 (bio, dis); Marín & Monserrat, 1987: 350-359 (bio, dis); Díaz-Aranda & Monserrat, 1988b: 227 (bio, dis), 1988c: 225 (bio, dis); Monserrat & Díaz-Aranda, 1988a: 94 (bio, dis), 1989: 55 (bio, dis), 1990: 47 (bio, dis); Meinander, 1990: 77 (rf); Marín & Monserrat, 1990: 228 (bio, dis), 1991b: 183 (bio, dis); Monserrat & Marín, 1992: 283 (bio); Plant, 1992b: 293 (rf); Marín, 1994: 260-297 (bio, dis, fen); Marín & Monserrat, 1995a: 45 (bio, dis), 1995b: 120 (bio, dis); Monserrat, 1995: 125 (bio, dis), 1996a: 15 (list); Blasco, 2000: 98 (bio, dis); H. Aspöck et al., 2001: 195 (rf); Monserrat, 2002: 15 (bio, dis), 2005a: 74 (bio, dis), 2013: 303 (rf) ; Monserrat & Triviño, 2013: 20 (list), 133 (dis); Letardi et al., 2013: 36 (list).
Especie conocida de Europa central y meridional (recientemente introducida en Gran Bretaña) y del N.O. de África. Generalmente
citada sobre coníferas: Cupressus, Thuja, Pinus, Cedrus, pero especialmente sobre Juniperus spp., y también sobre otros aciculifolios (Tetraclinis y Tamarix), y en menor número sobre Rosmarinus, Eucalyptus, Quercus y Pistacea, entre II-VIII, en altitudes de hasta 1920 m. En nuestra fauna tiene una clara tendencia térmica y mediterránea, incluyendo
Baleares (Formentera, Menorca), ha sido mayoritariamente citada a la luz y sobre coníferas (enebros, sabinas, cipreses, pinos,
alerces), puntualmente sobre encinas y alcornoques, entre II-X (sin duda con sucesión de generaciones), en altitudes que oscilan
entre 10-2.550 m.
Los estadios preimaginales de esta especie (Figs. 70-75) fueron descritos por Lauterbach (1972: 142) y Monserrat et al. (1991: 110).
NUEVO MATERIAL Y MATERIAL COMPLEMENTARIO ESTUDIADO: ESPAÑA: Ávila, Embalse de Burguillo, Barraco, 12.V.2007, 2♂♂, 2♀♀ sobre Juniperys oxycedrus, ♂♂, 3♀♀ sobre J. oxycedrus. Islas Baleares, Menorca, Cala Turqueta, 23.VII.2008, 1♂, 1♀ sobre J. phoenicea, F. Acevedo. Son Catlar, 23.VII.2008, 1♂, sobre Olea europaea, F. Acevedo. Cuenca, Vega del Codorno, 40°25’15.77’’N, 1°54’30.52’’O, 1480 m, 27.VI.2009, 1♂, 2♀♀ sobre J. sabina, 1♂ sobre J. oxycedrus. Granada, Hoya de la Mora, 30SVG6506, 2500 m, 30.VI.2011, 1♂ sobre Pinus sylvestris, La Herradura, Cerro Gordo, 7.IV.2007, 1♂, 3♀♀ a la luz, 2♂♂, 3♀♀ sobre J. oxycedrus, La Herradura, Punta de la Mona, 30SVF36, 200 m, 15.VI.2013, 1♂ a la luz. Guadalajara, Terzaga, 40°42′47”N, 1°55′01”O, 1347 m, 23.VII.2008, 1♀ sobre J. sabina. Huelva, Matalascañas, 20.VI.2012, 2♂♂, 5♀♀ sobre J. phoenicea. Jaén, Collado de los Jardines, 20.VI.2004, 1♀ sobre J. oxycedrus. Madrid, El Escorial, Arboreto Luis Ceballos, 7-21.VI.2008, 9♂♂, 2♀♀ en trampa de Malaise, 1.VIII-12.IX.2008, 1♂ en trampa de Malaise,
Madrid, 16.VI.2009, 1♂. Teruel, Bezas, 40°19′88′′N, 1°19′74′′O, 1163 m, 23.VII.2008, 1♂ sobre J. sabina, Laguna de Bezas, 6.VII.2010, 1♀ sobre J. thurifera, San Blas, 40°21′63”N, 1°10′03”O, 910 m, 22.VII.2008, 2♂♂, 1♀ sobre J. sabina (ver otro material en Helicoconis bachi n. sp.). ITALIA: Cerdeña, Sassari, Alghero, Capo Caccia, Dragunara, 40°34′32′′N, 8°09′39′′E, 10 m, 16.V.2008, 10♀♀ sobre J. phoenicea, Alghero, Fertilia, Monte Doglia, 40°36′37′′N, 8°14′43′′E, 190 m, 16.V.2008, 3♀♀ sobre Cupressus sempervirens, Sorso, Platamona, 40°49′10′′N, 8°31′02′′E, 5 m, 22.V.2008, 1♂, 2♀♀ sobre J. oxycedrus.
Semidalis vicina (Hagen, 1861) (Figs. 39, 76-78, 279-282)
Coniopteryx vicina Hagen, 1861
Hagen, 1861: 197 (d).
Semidalis vicina (Hagen, 1861)
Meinander, 1972a: 331 (rf); Gepp, 1974a: 224 (dis); Monserrat, 1978a: 65 (bio, dis), 1979a: 415 (bio, dis); Aspöck et al., 1980: 161 (bio, mf, tx, rf); Monserrat, 1980: 193 (bio, dis), 1984a: 149 (dis, tx), 1984b: 108 (bio, dis); Monserrat & Díaz-Aranda, 1987: 184 (bio, dis); Díaz-Aranda & Monserrat, 1988a: 121 (bio, dis), 1988c: 225 (bio, dis); Monserrat & Díaz-Aranda, 1990: 47 (bio, dis); Meinander, 1990: 79 (dis); Monserrat & Marín, 1992: 283 (bio); Monserrat et al., 1994: 60 (rf); Marín & Monserrat, 1995a: 45 (bio, dis); Monserrat, 1995: 124 (bio, dis); H. Aspöck et al., 2001: 196 (rf); Monserrat, 2002: 14 (bio, dis), 2005a: 74 (bio, dis); Letardi, 2012: 72 (dis); Monserrat, 2013: 303 (rf), 305 (rf); Monserrat & Triviño, 2013: 20 (list), 134 (dis); Letardi et al., 2013: 36 (list).
Semidalis ribesi Ohm, 1973
Meinander, 1972a: 330 (tax); Ohm, 1973a: 240 (d); Monserrat & Triviño, 2013: 20 (list).
Especie conocida del sur de Europa, Marruecos, Argelia y área central y oriental de Norteamérica (Canadá y USA). Asociada
a zonas térmicas y secas (pero no desérticas o subdesérticas) con condiciones climáticas mediterráneas, y recolectada sobre
una amplia variedad de sustratos arbóreos, sin especial predilección por alguno (tanto coníferas: Pinus, Juniperus, como planifolios y caducifolios: Quercus, Pistacea, Tamarix, Ceratonia, Castanea, Crataegus, Olea, Mirtus, Acer, Citrus, frutales, etc.), aunque probablemente no viva sobre ninguna de ellas y más prefiera vegetación arbustiva o baja, recolectada
entre IV-IX, en altitudes de 10 m hasta 1500 m. En nuestra fauna ha sido citada en medios mediterráneos, incluyendo Baleares
(Mallorca, Menorca), sobre encinas, coscojas, quejigos, endrinos, enebros, castaños, pistáceas, jaras, mirtos, entre V-X,
en altitudes que oscilan entre 10-1200 m.
Los estadios preimaginales de esta especie (Figs. 39, 76-78) fueron descritos o reiterados por Emerton (1906: 74), Muma (1967: 286, 1971: 284), Meinander (1974a: 14), Stange (1981: 1).
NUEVO MATERIAL ESTUDIADO: ESPAÑA: Almería, San José, 25.V.2003 1♀ a la luz. Cádiz, Chiclana, Pinar del Hierro, 20.VI.2012, 1♂ sobre Pistacea lentiscus, Grazalema, 31.V.2005, 770 m, 4 ej., A. Gruppe (AG). Parque Natural del Estrecho, 21.VI.2012, 1♂ sobre Olea europaea. Granada, La Herradura, Cantarriján, 30SVF36, 10 m, 12.VI.2012, 1♂ sobre Chamaerops humilis, V. Triviño, 30.VII.2013, 3♂♂ sobre P. lentiscus, La Taha, Pitres, 23.VII.2010, 1♂ sobre Castanea sativa. Huelva, La Nava, 19.VI.2012, 1♂, 1♀ sobre C. sativa. Málaga, Las Cañillas, 25.V.2012, 1♂, 2♀♀ sobre P. lentiscus.
ESPECIES INCORRECTA- O DUDOSAMENTE CITADAS EN LA FAUNA IBÉRICA:
Helicoconis lutea (Wallengren, 1871) (Figs. 20, 36, 42, 61-63, 79, 311)
Coniopteryx lutea Wallengren, 1871: 55 (d).
Especie holártica de carácter montano y extramediterráneo, conocida del centro y norte de Europa a Mongolia, norte de China
y Siberia hasta Península de Kamchatka (Vidlička, 2014) y está referida de Canadá (Alberta) y USA (Washington) en América del norte. Está citada como Coniopteryx lutea Wallengren, 1871 de forma muy imprecisa por Navás (1905b: 36): “Es de creer que... se encontrará en Castilla, pero no la he visto”. También hace referencia a ella (Navás, 1913a: 86) al describir A. laufferina, pero sin citar ningún ejemplar en concreto.
Refiriéndose a Helicoconis laufferina Navás, 1913 [sinónima de Helicoconis interna (Navás, 1911)], Ohm, 1965 comenta que algunos autores (Enderlein, 1930; Tjeder, 1957; Meinander, 1963) opinaron que podrían tratarse de sinónimos de esta especie (Helicoconis lutea), y consecuentemente formar parte de nuestra fauna, cosa que no ha sido así y hasta el presente no está citada en ella, donde
es poco probable exista, dada su distribución extramediterránea.
Estadios preimaginales en figuras 36, 42, 61-63, 79.
Coniopteryx (Coniopteryx) pygmaea Enderlein, 1906 (Figs. 26-30, 40, 170-172, 304)
Coniopteryx pygmaea Enderlein, 1906
Enderlein, 1906: 201 (d); Navás, 1915: 30 (dis), 1916a: 174 (dis), 1916b: 155 (dis), 1916c: 190 (dis); Bohigas & Sanchez, 1917: 318 (dis); Navás, 1917a: 42 (bio, dis), 1917b: 88 (dis), 1918c: 41 (dis), 1919a: 202 (dis), 1919b: 43 (dis), 1921a: 152 (dis), 1921b: 65 (dis), 1923b: 8 (dis), 1924a: 242, 244 (dis, key, mf), 1924b: 26 (dis), 1925a: 122, 123 (dis, key), 1925b: 136 (dis), 1928: 102 (dis), 1929a: 33 (dis), 1931: 121 (dis),1932: 16 (dis); Eglin, 1940: 270 (rf); Monserrat, 1977c: 142 (list), 1984d: 47 (rf); Marín & Monserrat, 1995b: 111 (rf); Monserrat & Triviño, 2013: 20 (list).
Coniopteryx (Coniopteryx) pygmaea Enderlein, 1906
Monserrat, 1984d: 47 (nt); Díaz-Aranda & Monserrat, 1988b: 226 (nt); Monserrat et al., 1994: 60 (rf); Monserrat, 1996a: 16 (list, tx); H. Aspöck et al., 2001: 185 (rf).
Según hemos argumentado con anterioridad, no aceptamos la sinonimia entre esta especie y C. parthenia propuesta por Günther (1993), por lo que nos mantenemos en el criterio de Enderlein (1906) y Meinander (1972a), no aceptando que ambas especies sean sinónimas, ni las referencias posteriores a nuestra fauna de autores que han asumido
esta sinonimia (Ejs.: H. Aspöck et al., 2001; Sziráki, 2004, 2011).
Especie conocida de centro Europa (según el criterio de Enderlein, 1906 y Meinander, 1972a) de Alemania, Austria, República Checa y Rumanía, asociada a planifolios en medios húmedos.
Como anteriormente hemos anotado, la posición taxonómica de esta especie desde su descripción (Enderlein, 1906: 201) ha sido bastante compleja y problemática, con multitud de datos bibliográficos de naturaleza faunística, morfológica
y biológica que parecían referirse a dos especies distintas, unas sobre planifolios, otras sobre coníferas. Ya hemos mencionado
que autores como Tjeder (1931), Killington (1936), Zelený (1961a), Kis (1964a), Kis et al. (1970) o Meinander (1962a) dibujaban como genitalia de C. pygmaea la que resultó ser genitalia de C. parthenia. Fue Meinander (1972a) quien puso fin a la mezcla de datos existentes entre esta especie y Coniopteryx parthenia, especie a la que asignó la información y citas dadas anteriormente sobre coníferas, restringiendo su distribución geográfica
al centro de Europa. Nuestra posición y criterio frente a otras opiniones porteriores ya ha sido anteriormente discutidos
y argumentados al tratar Coniopteryx parthenia, y con ello nos mantenemos el criterio de Enderlein (1906) y Meinander (1972a).
Como ocurrió en la fauna europea en general, la misma confusión se dio entre las citas antiguas de nuestra fauna, agravada
por la falta de adecuados criterios taxonómicos anotados anteriormente en la obra de L. Navás, quien, como anteriormente hemos
referido, la citó profusamente en nuestra fauna, generando consecuentemente posteriores referencias de esta especie en ella,
según hemos anotado.
C. pygmaea (sensu Meinander, 1972a) es una especie centroeuropea que, según nuestro criterio, hasta ahora no nos consta que viva en la Península Ibérica. Las citas de C. pygmaea en España anteriormemente anotadas fueron puestas en duda, discutidas o revisadas por Monserrat (1984d: 47) quien estudia los coniopterígidos de la colección personal de Navás y los ejemplares identificados y citados por él como C. pygmaea pertenecían, en realidad, a C. parthenia, C. haematica, C. esbenpeterseni, C. tjederi, C. lentiae, incluso a Semidalis aleyrodiformis y S. pseudouncinata, y Monserrat (1977c: 142), Díaz-Aranda & Monserrat (1988b: 226), Monserrat et al. (1994: 60), Marín & Monserrat (1995b: 111), Monserrat (1996a: 16) o Monserrat & Triviño (2013: 20) listan esta especie con ciertas/muchas dudas en base a las citas existentes en nuestra fauna, descartando su presencia
en la fauna ibérica. A pesar de que no existe ninguna cita aseverada de esta especie (C. pygmaea sensu Meinander, 1972a, nec Günther, 1993) en la fauna considerada, la hemos incluido en las claves para ilustrar datos de su morfología mencionados en el texto.
Estadios larvarios en Figs. 26-30, 40.
|
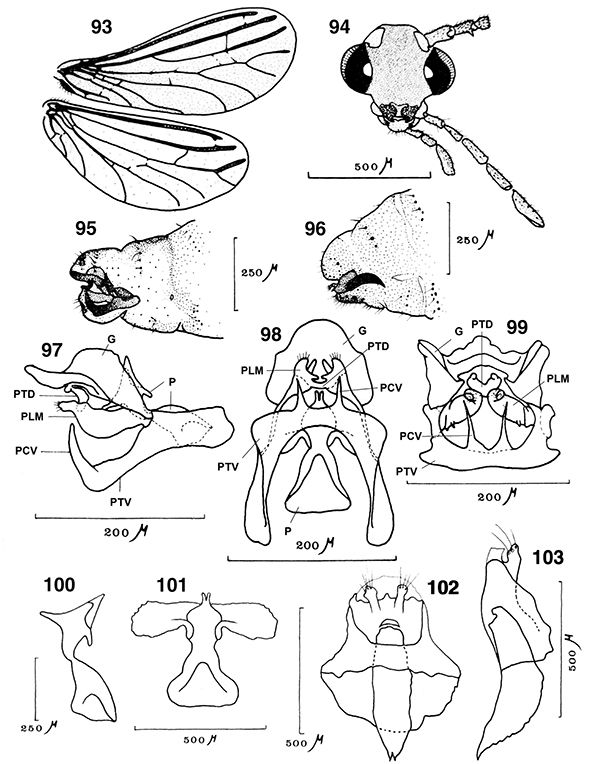 Figs. 93-103.— Aleuropteryx iberica, 93: alas, 94: cabeza, vista frontal, 95: extremo abdominal masculino, vista lateral, 96: ídem, femenino, 97: genitalia interna
masculina, vista lateral, 98: ídem, vista ventral, 99: ídem, vista caudal, 100: pene, vista lateral, 101: ídem, vista ventral,
102: bolsa copuladora, vista dorsal, 103: ídem, vista lateral (G: gonarco, P: pene, PCV: proceso caudo ventral, PLM: proceso
latero medial, PTD: placa tranversal dorsal, PTV: placa tranversal ventral), de Monserrat (1977b). Figs. 93-103.— Aleuropteryx iberica, 93: alas, 94: cabeza, vista frontal, 95: extremo abdominal masculino, vista lateral, 96: ídem, femenino, 97: genitalia interna
masculina, vista lateral, 98: ídem, vista ventral, 99: ídem, vista caudal, 100: pene, vista lateral, 101: ídem, vista ventral,
102: bolsa copuladora, vista dorsal, 103: ídem, vista lateral (G: gonarco, P: pene, PCV: proceso caudo ventral, PLM: proceso
latero medial, PTD: placa tranversal dorsal, PTV: placa tranversal ventral), de Monserrat (1977b).
Figs. 93-103.— Aleuropteryx iberica, 93: wings, 94: head, frontal view, 95: male abdominal tip, lateral view, 96: ditto, female, 97: male inner genitalia, lateral
view, 98: ditto, ventral view, 99: ditto, caudal view, 100: penis, lateral view, 101: ditto, ventral view, 102: bursa copulatrix,
dorsal view, 103: ditto, lateral view (G: gonarcus, P: penis, PCV: caudo ventral processus, PLM: latero medial processus,
PTD: tranversal dorsal plate, PTV: tranversal ventral plate), from Monserrat (1977b).
|
|
|
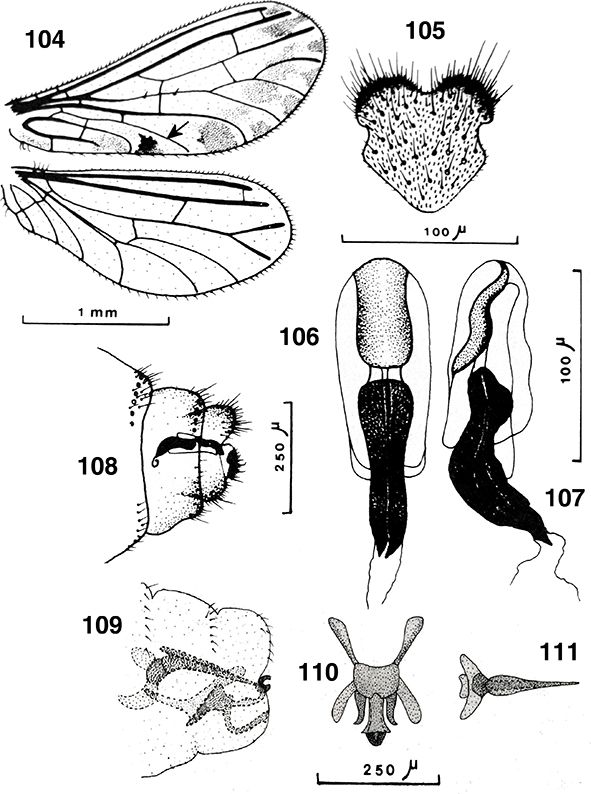 Figs. 104-111.— Aleuropteryx minuta, 104: alas, 105: placa subgenital femenina, vista ventral, 106: bolsa copuladora, vista dorso-anterior, 107: ídem, vista
lateral, 108: extremo abdominal femenino, vista lateral, 109: extremo abdominal masculino, vista lateral, 110: genitalia interna,
vista dorsal, 111: pene, vista dorsal. 104-108 de Monserrat (1985d). Figs. 104-111.— Aleuropteryx minuta, 104: alas, 105: placa subgenital femenina, vista ventral, 106: bolsa copuladora, vista dorso-anterior, 107: ídem, vista
lateral, 108: extremo abdominal femenino, vista lateral, 109: extremo abdominal masculino, vista lateral, 110: genitalia interna,
vista dorsal, 111: pene, vista dorsal. 104-108 de Monserrat (1985d).
Figs. 104-111.— Aleuropteryx minuta, 104: wings, 105: female subgenital plate, ventral view, 106: bursa copulatrix, dorso-anterior view, 107: ditto, lateral
view, 108: female abdominal tip, lateral view, 109: male abdominal tip, lateral view, 110: inner genitalia, dorsal view, 111:
penis, dorsal view. 104-108 from Monserrat (1985d).
|
|
|
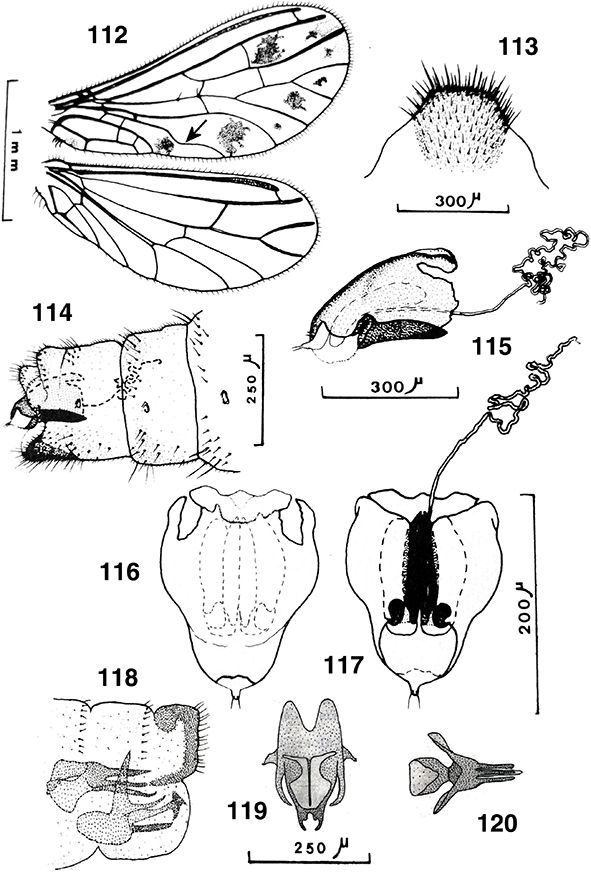 Figs. 112-120.— Aleuropteryx maculata, 112: alas, 113: placa subgenital femenina, vista ventral, 114: extremo abdominal femenino, vista lateral, 115: bolsa copuladora,
vista lateral, 116: ídem, vista dorsal, 117: ídem, vista ventral, 118: extremo abdominal masculino, vista lateral, 119: genitalia
interna, vista dorsal, 120: pene, vista dorsal. 112-117 de Monserrat (1984g). Figs. 112-120.— Aleuropteryx maculata, 112: alas, 113: placa subgenital femenina, vista ventral, 114: extremo abdominal femenino, vista lateral, 115: bolsa copuladora,
vista lateral, 116: ídem, vista dorsal, 117: ídem, vista ventral, 118: extremo abdominal masculino, vista lateral, 119: genitalia
interna, vista dorsal, 120: pene, vista dorsal. 112-117 de Monserrat (1984g).
Figs. 112-120.— Aleuropteryx maculata, 112: wings, 113 female subgenital plate, ventral view, 114: female abdominal tip, lateral view, 115: bursa copulatrix, lateral
view, 116: ditto, dorsal view, 117: ditto, ventral view, 118: male abdominal tip, lateral view, 119: inner genitalia, dorsal
view, 120: penis, dorsal view. 112-117 from Monserrat (1984g).
|
|
|
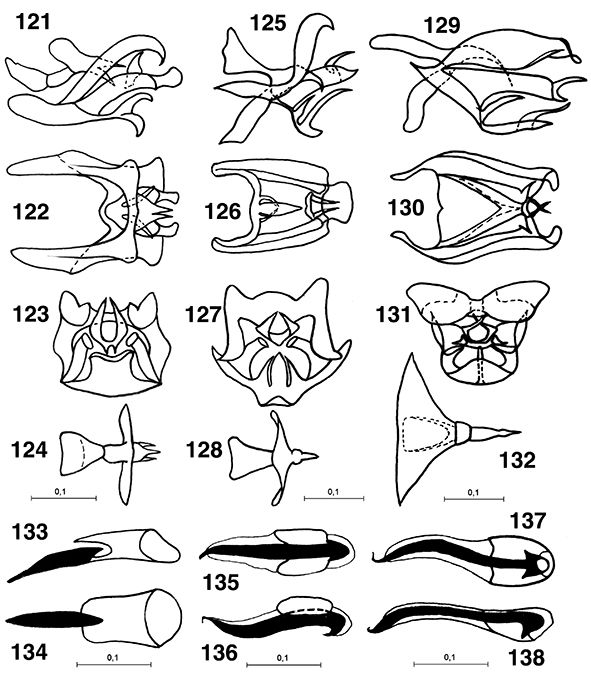 Figs. 121-138.— Aleuropteryx spp. 121-132: genitalia interna masculina y pene, vista lateral, genitalia interna, vista ventral y caudal, y pene, vista
dorsal de: 121-124: A. juniperi, 125-128: A. loewii, 129-132: A. remane. 133-138: bolsa copuladora femenina, vista dorsal y lateral de: 133-134: A. juniperi, 135-136: A. loewii, 137-138: A. remane. Escala en mm. Figs. 121-138.— Aleuropteryx spp. 121-132: genitalia interna masculina y pene, vista lateral, genitalia interna, vista ventral y caudal, y pene, vista
dorsal de: 121-124: A. juniperi, 125-128: A. loewii, 129-132: A. remane. 133-138: bolsa copuladora femenina, vista dorsal y lateral de: 133-134: A. juniperi, 135-136: A. loewii, 137-138: A. remane. Escala en mm.
Figs. 121-138.— Aleuropteryx spp. 121-132: male inner genitalia and penis, lateral view, inner genitalia, ventral and caudal view, and penis, dorsal view
of: 121-124: A. juniperi, 125-128: A. loewii, 129-132: A. remane. 133-138: female bursa copulatrix, dorsal and lateral view of: 133-134: A. juniperi, 135-136: A. loewii, 137-138: A. remane. Scale in mm.
|
|
|
 Figs. 139-158.— Genitalia masculina de Helicoconis spp. 139-144: H. bachi n. sp., 139: extremo abdominal, vista lateral, 140: ídem, vista ventral, 141: estilos, parámeros, pene, vista ventral, 142: estilo,
vista lateral, 143: pene, vista lateral, 144: parámero, vista lateral, 145-146: H. hirtinervis, 145: genitalia, vista ventral, 146: parámero, vista lateral, 147-149: H.hispanica, 147: genitalia, vista ventral, 148: parámero, vista lateral, 149: pene, vista lateral, 150-151: H. iberica, 150: pene, vista lateral, 151: estilo, vista ventral, 152-154: H. interna, 152: genitalia, vista ventral, 153: parámero, vista lateral, 154: pene, vista lateral, 155-157: H. panticosa, 155: parámero, vista lateral, 156: pene, vista lateral, 157: estilo, vista ventral, 158: H. bachi n. sp., estilo, vista ventral. Las flechas señalan algunos caracteres usados en las claves (ec: ectoprocto, hi: hipandrio, p: pene,
pa: parámero, pbe: proceso basal del estilo, pve: proceso ventral del ectoprocto, pvIX: proceso ventral del noveno segmento,
S: esternito, st: estilo). Fig. 155 de Ohm (1965). Escala en mm. Figs. 139-158.— Genitalia masculina de Helicoconis spp. 139-144: H. bachi n. sp., 139: extremo abdominal, vista lateral, 140: ídem, vista ventral, 141: estilos, parámeros, pene, vista ventral, 142: estilo,
vista lateral, 143: pene, vista lateral, 144: parámero, vista lateral, 145-146: H. hirtinervis, 145: genitalia, vista ventral, 146: parámero, vista lateral, 147-149: H.hispanica, 147: genitalia, vista ventral, 148: parámero, vista lateral, 149: pene, vista lateral, 150-151: H. iberica, 150: pene, vista lateral, 151: estilo, vista ventral, 152-154: H. interna, 152: genitalia, vista ventral, 153: parámero, vista lateral, 154: pene, vista lateral, 155-157: H. panticosa, 155: parámero, vista lateral, 156: pene, vista lateral, 157: estilo, vista ventral, 158: H. bachi n. sp., estilo, vista ventral. Las flechas señalan algunos caracteres usados en las claves (ec: ectoprocto, hi: hipandrio, p: pene,
pa: parámero, pbe: proceso basal del estilo, pve: proceso ventral del ectoprocto, pvIX: proceso ventral del noveno segmento,
S: esternito, st: estilo). Fig. 155 de Ohm (1965). Escala en mm.
Figs. 139-158.— Male genitalia of Helicoconis spp. 139-144: H. bachi n. sp., 139: abdominal apex, lateral view, 140: ditto, ventral view, 141: styli, paramera, penis, ventral view, 142: stylus, lateral
view, 143: penis, lateral view, 144: paramere, lateral view, 145-146: H. hirtinervis, 145: genitalia, ventral view, 146: paramere, lateral view, 147-149: H.hispanica, 147: genitalia, ventral view, 148: paramere, lateral view, 149: penis, lateral view, 150-151: H. iberica, 150: penis, lateral view, 151: stylus, ventral view. H. interna, 152: genitalia, ventral view, 153: paramere, lateral, 154: penis, lateral view, 155-157: H. panticosa, 155: paramere, lateral view, 156: penis, lateral view, 157: stylus, ventral view, 158: H. bachi n. sp., stylus, ventral view. Arrows indicate some characters used in the key (ec: ectoproct, hi: hypandrium, p: penis, pa: paramere,
pbe: basal processus of stylus, pve: ventral processus of ectoproct, pvIX: ventral processus of 9th segment, S: sternite,
st: stylus). Fig. 155 from Ohm (1965). Scale in mm.
|
|
|
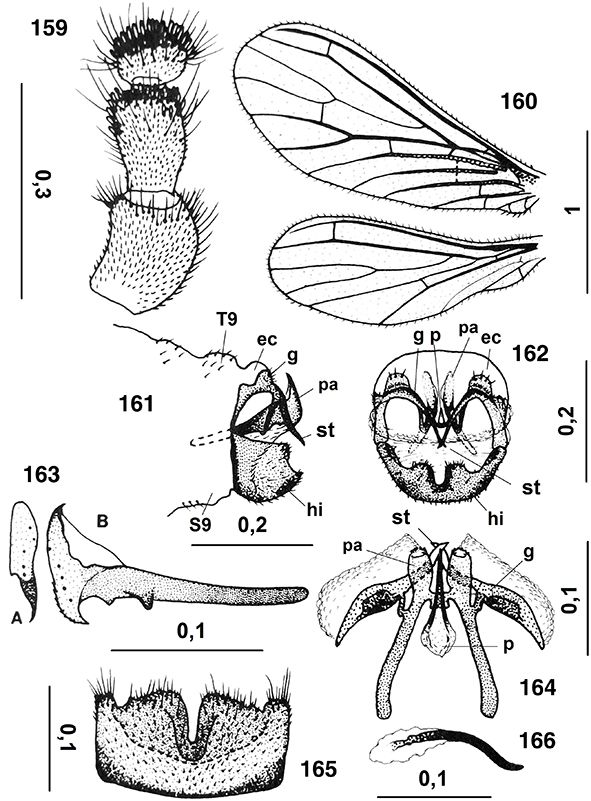 Figs. 159-166.— Coniopteryx (Coniopteryx) ezequi (♂), 159: primeros segmentos antenales, vista dorsal, 160: alas, 161: extremo abdominal, vista lateral, 162: ídem, vista
caudal, 163: parámero, A: vista caudal, B: vista lateral, 164: complejo gonarco-parámeros-pene, vista dorsal, 165: hipandrio,
vista ventral 166: pene, vista lateral (ec: ectoprocto, g: gonarco, hi: hipandrio, p: pene, pa: parámero, S: esternito, st:
estilo, T: terguito). Escala en mm. Adaptado de Monserrat (1984g). Figs. 159-166.— Coniopteryx (Coniopteryx) ezequi (♂), 159: primeros segmentos antenales, vista dorsal, 160: alas, 161: extremo abdominal, vista lateral, 162: ídem, vista
caudal, 163: parámero, A: vista caudal, B: vista lateral, 164: complejo gonarco-parámeros-pene, vista dorsal, 165: hipandrio,
vista ventral 166: pene, vista lateral (ec: ectoprocto, g: gonarco, hi: hipandrio, p: pene, pa: parámero, S: esternito, st:
estilo, T: terguito). Escala en mm. Adaptado de Monserrat (1984g).
Figs. 159-166.— Coniopteryx (Coniopteryx) ezequi (♂), 159: basal antennal segments, dorsal view, 160: wings, 161: abdominal tip, lateral view, 162: ditto, caudal view, 163:
paramere, A: caudal view, B: lateral view, 164: gonarcus-paramera-penis complex, dorsal view, 165: hypandrium, ventral view,
166: penis, lateral view (ec: ectoproct, g: gonarcus, hi: hypandrium, p: penis, pa: paramere, S: sternite, st: stylus, T:
tergite). Scale in mm. Adapted from Monserrat (1984g).
|
|
|
 Figs. 167-178.— Genitalia masculina de Coniopteryx (Coniopteryx) spp., extremo abdominal, vista lateral, parámero, vista lateral e hipandrio, vista ventral de: 167-169: C. (C.) parthenia, (sensu Meinander, 1972a), 170-172: C. (C.) pygmaea, 173-175: C. (C.) tineiformis, 176-178: C. (C.) borealis (ec: ectoprocto, g: gonarco, hi: hipandrio, p: parámero, S: esternito, st: estilo, T: terguito). Escala en mm. Figs. 167-178.— Genitalia masculina de Coniopteryx (Coniopteryx) spp., extremo abdominal, vista lateral, parámero, vista lateral e hipandrio, vista ventral de: 167-169: C. (C.) parthenia, (sensu Meinander, 1972a), 170-172: C. (C.) pygmaea, 173-175: C. (C.) tineiformis, 176-178: C. (C.) borealis (ec: ectoprocto, g: gonarco, hi: hipandrio, p: parámero, S: esternito, st: estilo, T: terguito). Escala en mm.
Figs. 167-178.— Male Genitalia of Coniopteryx (Coniopteryx) spp., abdominal apex, lateral view, paramere, lateral view and hypandrium, ventral view in: 167-169: C. (C.) parthenia (sensu Meinander, 1972a), 170-172: C. (C.) pygmaea, 173-175: C. (C.) tineiformis, 176-178: C. (C.) borealis (ec: ectoproct, g: gonarcus, hi: hypandrium, p: paramere, S: sternite, st: stylus, T: tergite). Scale in mm.
|
|
|
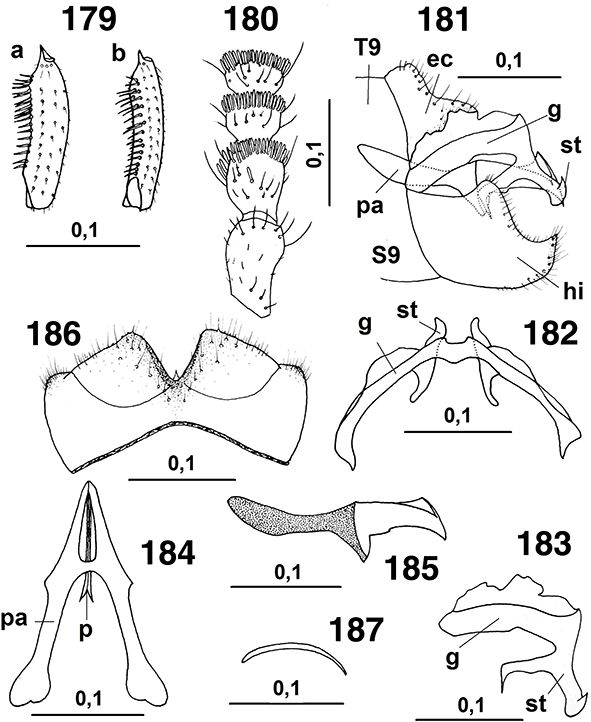 Figs. 179-187.— Coniopteryx (Holoconiopteryx) drammonti (♂), 179: fémur anterior, a: vista lateral, b: vista antero-lateral, 180: primeros segmentos antenales, vista dorsal, 181: genitalia, vista lateral, 182: gonarco-estilos, vista dorsal, 183: ídem, vista lateral, 184: parámeros-pene, vista dorsal, 185: parámero, vista lateral, 186: hipandrio, vista ventral, 187: pene, vista lateral (ec: ectoprocto, g: gonarco, hi: hipandrio, p: pene, pa: parámero, S: esternito, st: estilo, T: terguito). Escala en mm. Adaptado de Monserrat (1975). Figs. 179-187.— Coniopteryx (Holoconiopteryx) drammonti (♂), 179: fémur anterior, a: vista lateral, b: vista antero-lateral, 180: primeros segmentos antenales, vista dorsal, 181: genitalia, vista lateral, 182: gonarco-estilos, vista dorsal, 183: ídem, vista lateral, 184: parámeros-pene, vista dorsal, 185: parámero, vista lateral, 186: hipandrio, vista ventral, 187: pene, vista lateral (ec: ectoprocto, g: gonarco, hi: hipandrio, p: pene, pa: parámero, S: esternito, st: estilo, T: terguito). Escala en mm. Adaptado de Monserrat (1975).
Figs. 179-187.— Coniopteryx (Holoconiopteryx) drammonti (♂), 179: anterior femur, a: lateral view, b: antero-lateral view, 180: basal antennal segments, 181: genitalia, lateral view, 182: gonarcus-stylus, dorsal view, 183: ditto, lateral view, 184: paramera-penis, dorsal view, 185: paramere, lateral view, 186: hypandrium, ventral view, 187: penis, lateral view (ec: ectoproct, g: gonarcus, hi: hypandrium, p: penis, pa: paramere, S: sternite, st: stylus, T: tergite). Scale in mm. Adapted from Monserrat (1975).
|
|
|
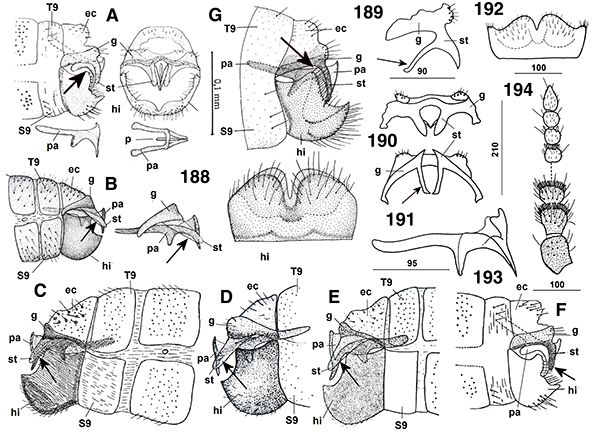 Figs. 188-194.— Coniopteryx (Holoconiopteryx) spp. 188: C. (H.) tullgreni, sp. reval. genitalia masculina adaptado según varios autores: A: de Meinander (1972a), B: de Kis et al. (1970), C: de Tjeder (1931), D: de Kis (1964a), E: de Meinander (1962a), F: de Dorokhova (1987), G: de Aspöck et al. (1980), 189-194: Coniopteryx (H.) haematica (♂): 189: gonarco y estilo, vista lateral, 190: ídem, vista caudal, 191: ídem, vista ventral, 192: hipandrio, vista ventral,
193: parámero, vista lateral, 194: segmentos basales y terminales de la antena. Las flechas señalan algunos caracteres usados
en el texto (ec: ectoprocto, g: gonarco, hi: hipandrio, p: pene, pa: parámero, S: esternito, st: estilo, T: terguito). 189-194:
escala en μm, adaptado de Monserrat (1981c). Figs. 188-194.— Coniopteryx (Holoconiopteryx) spp. 188: C. (H.) tullgreni, sp. reval. genitalia masculina adaptado según varios autores: A: de Meinander (1972a), B: de Kis et al. (1970), C: de Tjeder (1931), D: de Kis (1964a), E: de Meinander (1962a), F: de Dorokhova (1987), G: de Aspöck et al. (1980), 189-194: Coniopteryx (H.) haematica (♂): 189: gonarco y estilo, vista lateral, 190: ídem, vista caudal, 191: ídem, vista ventral, 192: hipandrio, vista ventral,
193: parámero, vista lateral, 194: segmentos basales y terminales de la antena. Las flechas señalan algunos caracteres usados
en el texto (ec: ectoprocto, g: gonarco, hi: hipandrio, p: pene, pa: parámero, S: esternito, st: estilo, T: terguito). 189-194:
escala en μm, adaptado de Monserrat (1981c).
Figs. 188-194.— Coniopteryx (Holoconiopteryx) spp. 188: C. (H.) tullgreni sp. reval. adapted from autors: A: by Meinander (1972a), B: by Kis et al. (1970), C: by Tjeder (1931), D: by Kis (1964a), E: by Meinander (1962a), F: by Dorokhova (1987), G: by Aspöck et al. (1980), 189-194: Coniopteryx (H.) haematica (♂): 189: gonarcus and stylus, lateral view, 190: ditto, caudal view, 191: ditto, ventral view, 192: hypandrium, ventral
view, 193: paramere, lateral view, 194: basal and distal antennal segments. Arrows indicate some characters used in the text
(ec: ectoproct, g: gonarcus, hi: hypandrium, p: penis, pa: paramere, S: sternite, st: stylus, T: tergite). 189-194: scale
in μm, adapted from Monserrat (1981c).
|
|
|
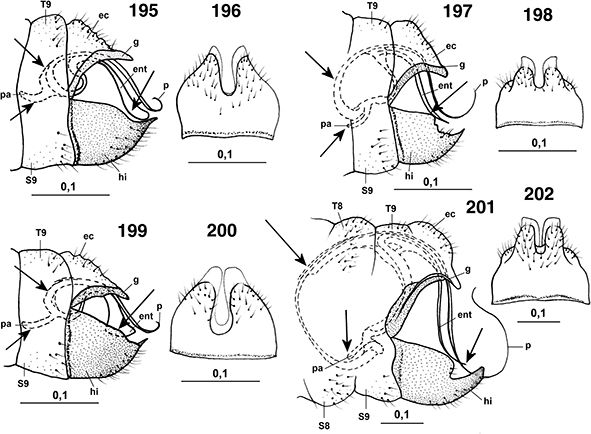 Figs. 195-202.— Genitalia masculina de Coniopteryx (Metaconiopteryx) spp., extremo abdominal, vista lateral e hipandrio, vista ventral de: 195, 196: C. (M.) esbenpeterseni, 197, 198: C. (M.) lentiae, 199, 200: C. (M.) arcuata, 201, 202: C. (M.) tjederi (ec: ectoprocto, ent: entoproceso, g: gonarco, hi: hipandrio, p: pene, pa: parámero, S: esternito, T: terguito). Las flechas
señalan algunos caracteres usados en las claves. Escala en mm. Figs. 195-202.— Genitalia masculina de Coniopteryx (Metaconiopteryx) spp., extremo abdominal, vista lateral e hipandrio, vista ventral de: 195, 196: C. (M.) esbenpeterseni, 197, 198: C. (M.) lentiae, 199, 200: C. (M.) arcuata, 201, 202: C. (M.) tjederi (ec: ectoprocto, ent: entoproceso, g: gonarco, hi: hipandrio, p: pene, pa: parámero, S: esternito, T: terguito). Las flechas
señalan algunos caracteres usados en las claves. Escala en mm.
Figs. 195-202.— Male genitalia of Coniopteryx (Metaconiopteryx) spp., abdominal apex, lateral view and hypandrium, ventral view in: 195, 196: C. (M.) esbenpeterseni, 197, 198: C. (M.) lentiae, 199, 200: C. (M.) arcuata, 201, 202: C. (M.) tjederi (ec: ectoproct, ent: entoprocessus, g: gonarcus, hi: hypandrium, p: penis, pa: paramere, S: sternite, T: tergite). Harrows
indicates some characters used in keys. Scale in mm.
|
|
|
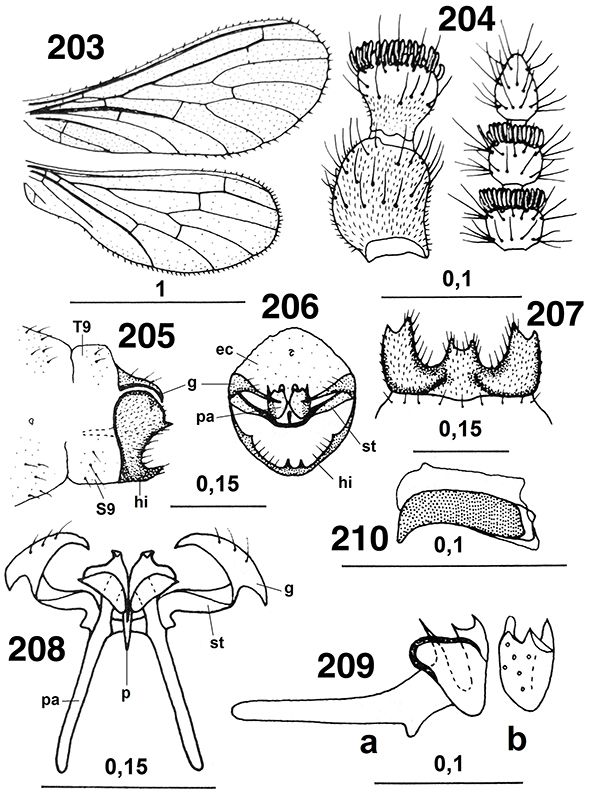 Figs. 203-210.— Coniopteryx (Xeroconiopteryx) atlantica (♂), 203: alas, 204: escapo-pedicelo y extremo antenal, 205: extremo abdominal, vista lateral, 206: ídem, vista caudal, 207:
hipandrio, vista ventral, 208: complejo gonarco-estilo-parámeros-pene, vista dorsal, 209: parámero, a: vista lateral-interna,
b: vista caudal, 210: pene, vista lateral (ec: ectoprocto, g: gonarco, hi: hipandrio, p: pene, pa: parámero, S: esternito,
st: estilo, T: terguito). Escala en mm. Adaptado de Monserrat (1985d). Figs. 203-210.— Coniopteryx (Xeroconiopteryx) atlantica (♂), 203: alas, 204: escapo-pedicelo y extremo antenal, 205: extremo abdominal, vista lateral, 206: ídem, vista caudal, 207:
hipandrio, vista ventral, 208: complejo gonarco-estilo-parámeros-pene, vista dorsal, 209: parámero, a: vista lateral-interna,
b: vista caudal, 210: pene, vista lateral (ec: ectoprocto, g: gonarco, hi: hipandrio, p: pene, pa: parámero, S: esternito,
st: estilo, T: terguito). Escala en mm. Adaptado de Monserrat (1985d).
Figs. 203-210.— Coniopteryx (Xeroconiopteryx) atlantica (♂), 203: wings, 204: scape-pedicellus and antennal tip, 205: abdominal tip, lateral view, 206: ditto, caudal view, 207:
hypandrum, ventral view, 208: gonarcus-stylus-paramera-penis, dorsal view, 209: paramere, a: inner-lateral view, b: caudal
view, 210: penis, lateral view (ec: ectoproct, g: gonarcus, hi: hypandrium, p: penis, pa: paramere, S: sternite, st: stylus,
T: tergite). Scale in mm. Adapted from Monserrat (1985d).
|
|
|
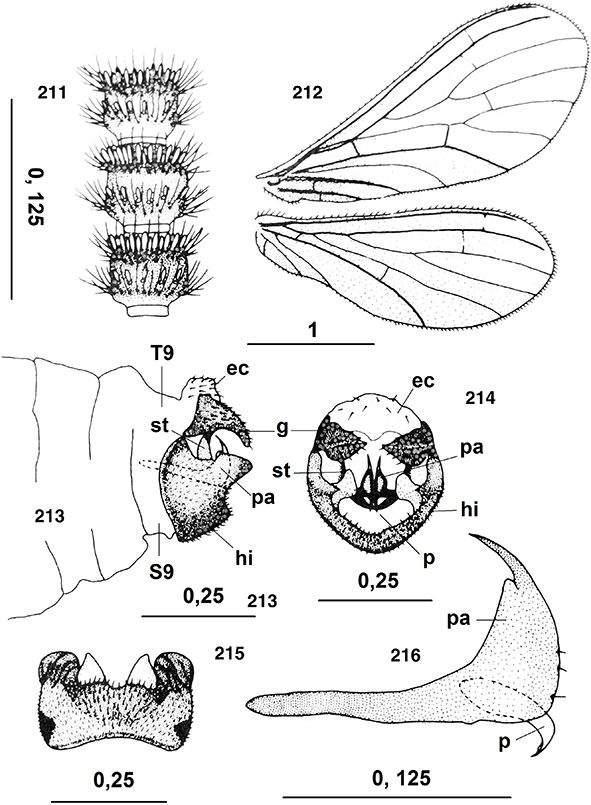 Figs. 211-216.— Coniopteryx (Xeroconiopteryx) kerzhneri (♂), 211: primeros segmentos del flagelo antenal, 212: alas, 213: extremo abdominal, vista lateral, 214: ídem, vista caudal,
215: hipandrio, vista ventral, 216: parámero y pene, vista lateral (ec: ectoprocto, g: gonarco, hi: hipandrio, p: pene, pa:
parámero, S: esternito, st: estilo, T: terguito). Escala en mm. Adaptado de Monserrat (1982b). Figs. 211-216.— Coniopteryx (Xeroconiopteryx) kerzhneri (♂), 211: primeros segmentos del flagelo antenal, 212: alas, 213: extremo abdominal, vista lateral, 214: ídem, vista caudal,
215: hipandrio, vista ventral, 216: parámero y pene, vista lateral (ec: ectoprocto, g: gonarco, hi: hipandrio, p: pene, pa:
parámero, S: esternito, st: estilo, T: terguito). Escala en mm. Adaptado de Monserrat (1982b).
Figs. 211-216.— Coniopteryx (Xeroconiopteryx) kerzhneri (♂), 211: basal flagelomeres, 212: wings, 213: abdominal tip, lateral view, 214: ditto, caudal view, 215: hypandrium, ventral
view, 216: paramere and penis, lateral view (ec: ectoproct, g: gonarcus, hi: hypandrium, p: penis, pa: paramere, S: sternite,
st: stylus, T: tergite). Scale in mm. Adapted from Monserrat (1982b).
|
|
|
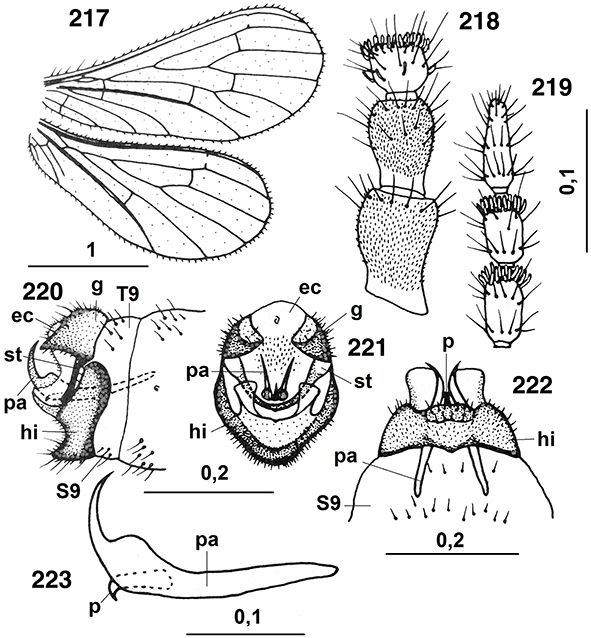 Figs. 217-223.— Coniopteryx (Xeroconiopteryx) ketiae (♂), 217: alas, 218: segmentos basales de la antena, 219: extremo antenal, 220: extremo abdominal, vista lateral, 221: ídem,
vista caudal, 222: ídem, vista ventral, 223: parámero-pene, vista lateral (ec: ectoprocto, g: gonarco, hi: hipandrio, p: pene,
pa: parámero, S: esternito, st: estilo, T: terguito). Escala en mm. Adaptado de Monserrat (1985d). Figs. 217-223.— Coniopteryx (Xeroconiopteryx) ketiae (♂), 217: alas, 218: segmentos basales de la antena, 219: extremo antenal, 220: extremo abdominal, vista lateral, 221: ídem,
vista caudal, 222: ídem, vista ventral, 223: parámero-pene, vista lateral (ec: ectoprocto, g: gonarco, hi: hipandrio, p: pene,
pa: parámero, S: esternito, st: estilo, T: terguito). Escala en mm. Adaptado de Monserrat (1985d).
Figs. 217-223.— Coniopteryx (Xeroconiopteryx) ketiae (♂), 217: wings, 218: basal antennal segments, 219: antennal tip, 220: abdominal tip, view lateral, 221: ditto, caudal view,
222: ditto, ventral view, 223: paramere-penis, lateral view (ec: ectoproct, g: gonarcus, hi: hypandrium, p: penis, pa: paramere,
S: sternite, st: stylus, T: tergite). Scale in mm. Adapted from Monserrat (1985d).
|
|
|
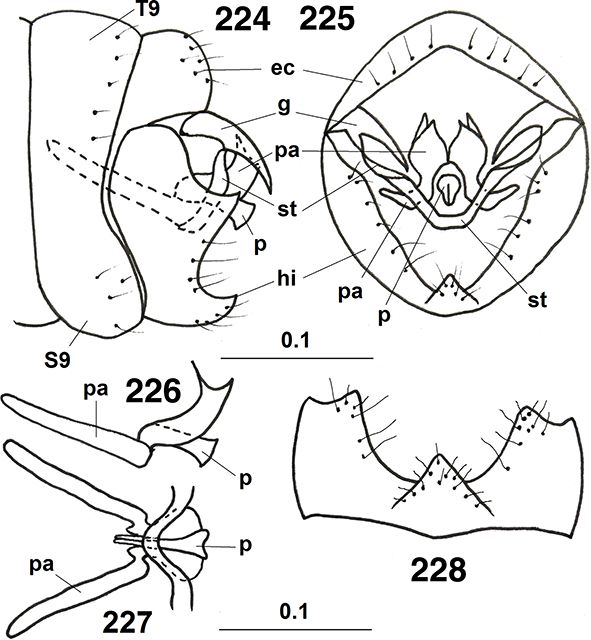 Figs. 224-228.— Coniopteryx (Xeroconiopteryx) loipetsederi (♂), 224: extremo abdominal, vista lateral, 225: ídem, vista caudal, 226: parámero-pene, vista lateral, 227: ídem, vista
ventral, 228: hipandrio, vista ventral (ec: ectoprocto, g: gonarco, hi: hipandrio, p: pene, pa: parámero, S: esternito, st:
estilo, T: terguito). Escala en mm. Figs. 224-228.— Coniopteryx (Xeroconiopteryx) loipetsederi (♂), 224: extremo abdominal, vista lateral, 225: ídem, vista caudal, 226: parámero-pene, vista lateral, 227: ídem, vista
ventral, 228: hipandrio, vista ventral (ec: ectoprocto, g: gonarco, hi: hipandrio, p: pene, pa: parámero, S: esternito, st:
estilo, T: terguito). Escala en mm.
Figs. 224-228.— Coniopteryx (Xeroconiopteryx) loipetsederi (♂), 224: abdominal tip, view lateral, 225: ditto, caudal view, 226: paramere-penis, lateral view, 227: ditto, ventral view,
228: hypandrium, ventral view (ec: ectoproct, g: gonarcus, hi: hypandrium, p: penis, pa: paramere, S: sternite, st: stylus,
T: tergite). Scale in mm.
|
|
|
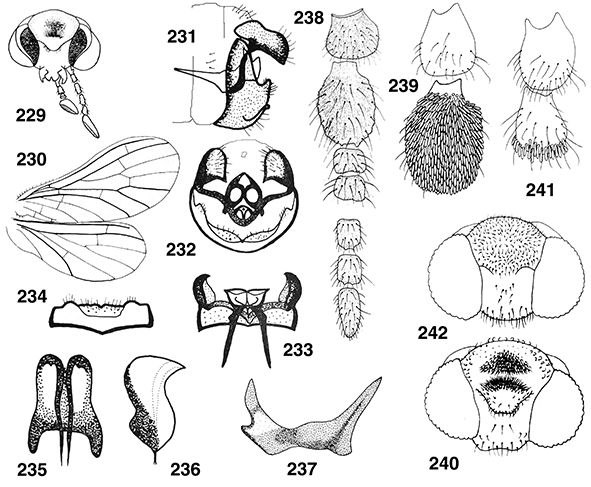 Figs. 229-242.— Coniopteryx (Xeroconiopteryx) perisi (♂), 229: cabeza, vista frontal, 230: alas, 231: extremo abdominal, vista lateral, 232: ídem, vista caudal, 233: ídem, vista
dorsal, 234: hipandrio, vista ventral, 235: pene, vista ventral, 236: ídem, vista lateral, 237: parámero, vista lateral, 238:
primeros y últimos segmentos antenales, adaptado de Monserrat (1976a), 239: quetotaxia del escapo y pedicelo, 240: esquema de la cabeza masculina mostrando sus elementos sobre el vértex, de Monserrat (2002) (ec: ectoprocto, g: gonarco, hi: hipandrio, p: pene, pa: parámero, S: esternito, st: estilo, T: terguito). 241-242: Coniopteryx (Xeroconiopteryx) atlasensis (♂), 241: quetotaxia del escapo y pedicelo, 242: esquema de la cabeza masculina mostrando sus elementos sobre el vértex, de Monserrat (2002). A diferentes escalas. Figs. 229-242.— Coniopteryx (Xeroconiopteryx) perisi (♂), 229: cabeza, vista frontal, 230: alas, 231: extremo abdominal, vista lateral, 232: ídem, vista caudal, 233: ídem, vista
dorsal, 234: hipandrio, vista ventral, 235: pene, vista ventral, 236: ídem, vista lateral, 237: parámero, vista lateral, 238:
primeros y últimos segmentos antenales, adaptado de Monserrat (1976a), 239: quetotaxia del escapo y pedicelo, 240: esquema de la cabeza masculina mostrando sus elementos sobre el vértex, de Monserrat (2002) (ec: ectoprocto, g: gonarco, hi: hipandrio, p: pene, pa: parámero, S: esternito, st: estilo, T: terguito). 241-242: Coniopteryx (Xeroconiopteryx) atlasensis (♂), 241: quetotaxia del escapo y pedicelo, 242: esquema de la cabeza masculina mostrando sus elementos sobre el vértex, de Monserrat (2002). A diferentes escalas.
Figs. 229-242.— Coniopteryx (Xeroconiopteryx) perisi (♂), 229: head, frontal view, 230: wings, 231: abdominal apex, lateral view, 232: ditto, caudal view, 233: ditto, dorsal
view, 234: hypandrium, ventral view, 235: penis, ventral view, 236: ditto, lateral view, 237: paramere, lateral view, 238:
basal and distal antennal segments, adapted from Monserrat (1976a), 239: chaetotaxy of scape and pedicellus, 240: scheme of male head showing its elements on vertex, from Monserrat (2002) (ec: ectoproct, g: gonarcus, hi: hypandrium, p: penis, pa: paramere, S: sternite, st: stylus, T: tergite). 241-242: Coniopteryx (Xeroconiopteryx) atlasensis (♂), 241: chaetotaxy of scape and pedicellus, 242: scheme of male head showing its elements on vertex, from Monserrat (2002). At diferent scales.
|
|
|
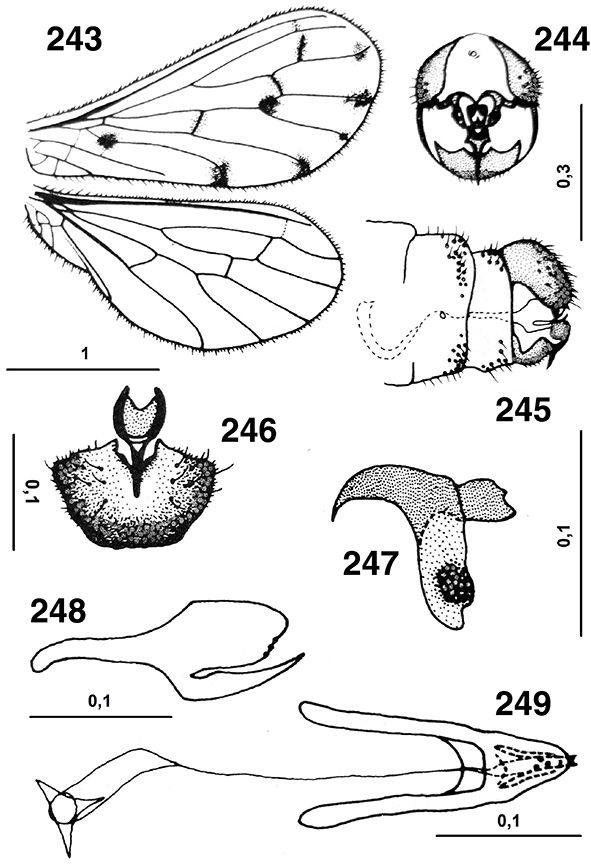 Figs. 243-249.— Nimboa adelae (♂), 243: alas, 244: extremo abdominal, vista caudal, 245: ídem, vista lateral, 246: hipandrio-pene, vista ventral, 247:
gonarco, vista lateral externa, 248: parámeros, vista lateral, 249: ídem, vista dorsal. Escala en mm. De Monserrat (1985d). Figs. 243-249.— Nimboa adelae (♂), 243: alas, 244: extremo abdominal, vista caudal, 245: ídem, vista lateral, 246: hipandrio-pene, vista ventral, 247:
gonarco, vista lateral externa, 248: parámeros, vista lateral, 249: ídem, vista dorsal. Escala en mm. De Monserrat (1985d).
Figs. 243-249.— Nimboa adelae (♂), 243: wings, 244: abdominal apex, caudal view, 245: ditto, lateral view, 246: hypandrium-penis, ventral view, 247: gonarcus,
latero-external view, 248: paramera, lateral view, 249: ditto, dorsal view. Scale in mm. From Monserrat (1985d).
|
|
|
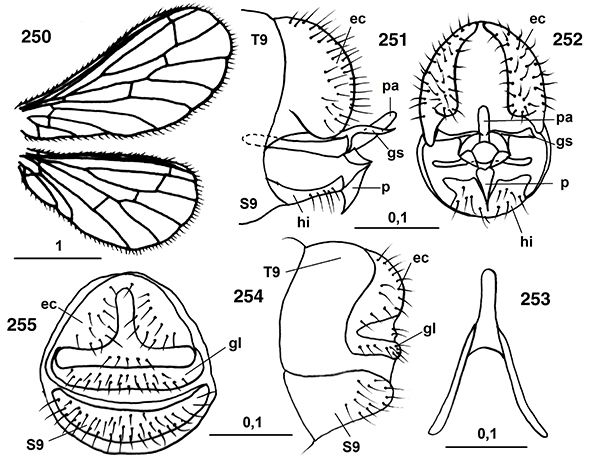 Figs. 250-255.— Nimboa espanoli, 250: alas, 251: extremo abdominal masculino, vista lateral, 252: ídem, vista caudal, 253: parámeros, vista ventral, 254:
extremo abdominal femenino, vista latreral, 255: ídem, vista caudal (ec: ectoprocto, gl: gonapófisis laterales, gs: gonarco,
hi: hipandrio, p: pene, pa: parámeros, S: esternito, T: terguito). Escala en mm. Figs. 250-255.— Nimboa espanoli, 250: alas, 251: extremo abdominal masculino, vista lateral, 252: ídem, vista caudal, 253: parámeros, vista ventral, 254:
extremo abdominal femenino, vista latreral, 255: ídem, vista caudal (ec: ectoprocto, gl: gonapófisis laterales, gs: gonarco,
hi: hipandrio, p: pene, pa: parámeros, S: esternito, T: terguito). Escala en mm.
Figs. 250-255.— Nimboa espanoli, 250: wings, 251: male abdominal apex, lateral view, 252: ditto, caudal view, 253: paramera, ventral view, 254: female abdominal
apex, lateral view, 255: ditto, caudal view (ec: ectoproct, gl: lateral gonapophysis, gs: gonarcus, hi: hypandrium, p: penis,
pa: paramera, S: sternite, T: tergite). Scale in mm.
|
|
|
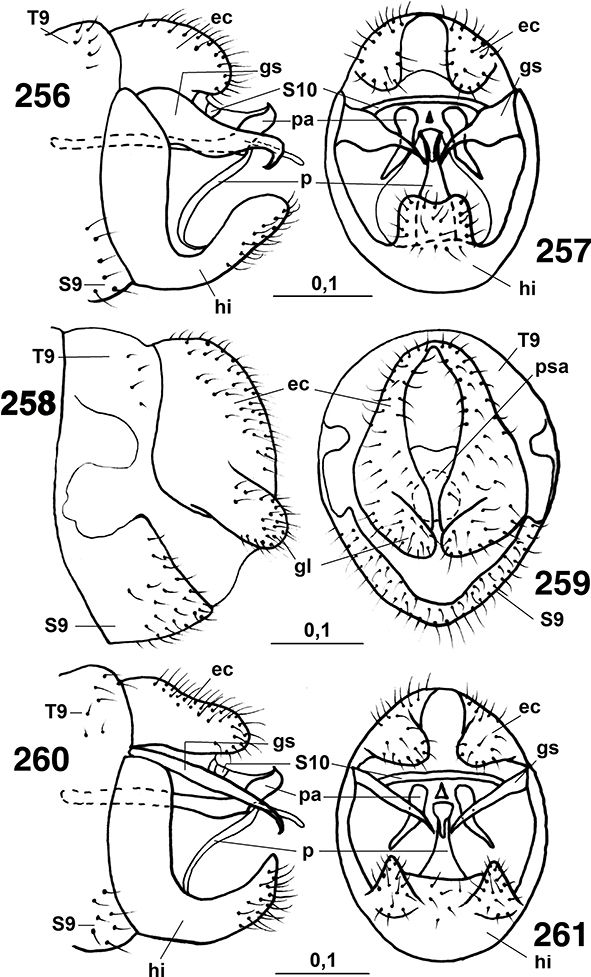 Figs. 256-261.— Extremo abdominal de Parasemidalis spp. 256-259: P. fuscipennis, 256: extremo abdominal masculino, vista lateral, 257: ídem, vista caudal, 258: extremo abdominal femenino, vista lateral,
259: ídem, vista caudal, 260-261: P. triton, 260: extremo abdominal masculino, vista lateral, 261: ídem, vista caudal (ec: ectoprocto, gl: gonapófisis laterales, gs:
gonarco, hi: hipandrio, p: pene, pa: parámero, psa: placa subanal, S: esternito, T: terguito). Escala en mm. Figs. 256-261.— Extremo abdominal de Parasemidalis spp. 256-259: P. fuscipennis, 256: extremo abdominal masculino, vista lateral, 257: ídem, vista caudal, 258: extremo abdominal femenino, vista lateral,
259: ídem, vista caudal, 260-261: P. triton, 260: extremo abdominal masculino, vista lateral, 261: ídem, vista caudal (ec: ectoprocto, gl: gonapófisis laterales, gs:
gonarco, hi: hipandrio, p: pene, pa: parámero, psa: placa subanal, S: esternito, T: terguito). Escala en mm.
Figs. 256-261.— Abdominal apex in Parasemidalis spp. 256-259: P. fuscipennis, 256: male abdominal tip, lateral view, 257: ditto, caudal view, 258: female abdominal tip, lateral view, 259: ditto, caudal
view, 260-261: P. triton, 260: male abdominal tip, lateral view, 261: ditto, caudal view (ec: ectoproct, gl: lateral gonapophysis, gs: gonarcus, hi:
hypandrium, p: penis, pa: paramere, psa: subanal plate, S: sternite, T: tergite). Scale in mm.
|
|
|
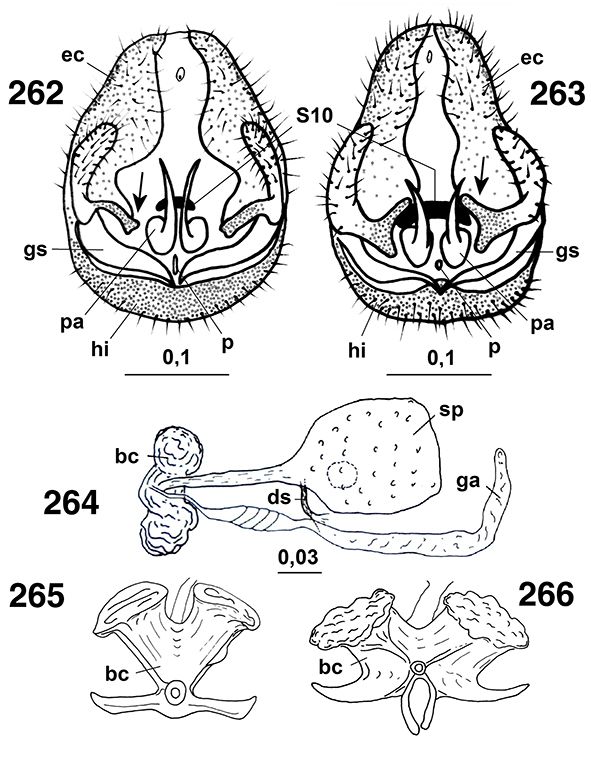 Figs. 262-266.— Genitalia externa e interna de Conwentzia spp. 262: extremo abdominal masculino, vista caudal de C. psociformis, 263: ídem de C. pineticola. Las flechas señalan algunos caracteres usados en las claves. Escala en mm. 264: estructuras genitales internas en la hembra
de C. psociformis, vista dorsal. Escala en mm. Adaptado de Sziráki (1996), 265: bolsa copuladora de C. psociformis, vista ventral, 266: ídem de C. pineticola, vista ventral. A diferentes escalas. Adaptado de Sziráki (1992a) (bc: bolsa copuladora, ds: ducto seminal, ec: ectoprocto, ga: glándula accesoria, gs: gonarco, hi: hipandrio, p: pene, pa:
parámero, S: esternito, sp: espermateca). Figs. 262-266.— Genitalia externa e interna de Conwentzia spp. 262: extremo abdominal masculino, vista caudal de C. psociformis, 263: ídem de C. pineticola. Las flechas señalan algunos caracteres usados en las claves. Escala en mm. 264: estructuras genitales internas en la hembra
de C. psociformis, vista dorsal. Escala en mm. Adaptado de Sziráki (1996), 265: bolsa copuladora de C. psociformis, vista ventral, 266: ídem de C. pineticola, vista ventral. A diferentes escalas. Adaptado de Sziráki (1992a) (bc: bolsa copuladora, ds: ducto seminal, ec: ectoprocto, ga: glándula accesoria, gs: gonarco, hi: hipandrio, p: pene, pa:
parámero, S: esternito, sp: espermateca).
Figs. 262-266.— External and internal genitalia in Conwentzia spp. 262: Male abdominal tip, cuadal view in C. psociformis, 263: ditto of C. pineticola. Harrows indicates some characters used in keys. Scale in mm. 264: female internal structures in C. psociformis, dorsal view. Scale in mm. Adapted from Sziráki (1996), 265: bursa copulatrix in C. psociformis, ventral view, 266: ditto in C. pineticola, ventral view. At different scales. Adapted from Sziráki (1992a) (bc: bursa copulatrix, ds: ductus seminalis, ec: ectoproct, ga: accessory gland, g: gonarcus, hi: hypandrium, p: penis,
pa: paramera, S: sternite, sp: spermatheca).
|
|
|
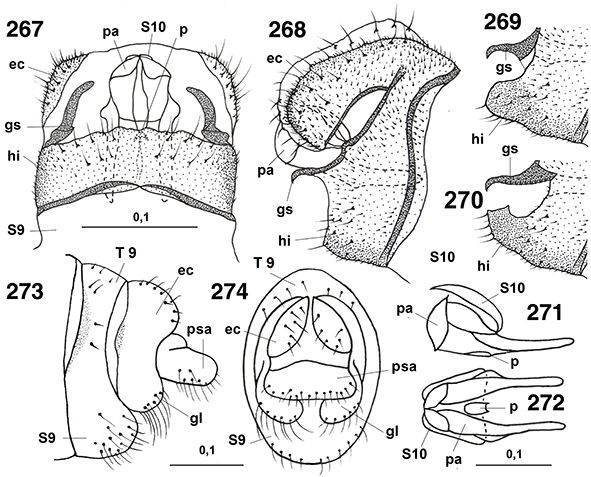 Figs. 267-274.— Hemisemidalis pallida, 267: extremo abdominal masculino, vista ventral, 268: ídem, vista lateral (adaptado de Monserrat, 2010), 269-270: variabilidad en el margen caudal del hipandrio y del gonarco, 271: complejo parámeros-pene-décimo esternito, vista
lateral, 272: ídem, vista ventral, 273: extremo abdominal femenino, vista lateral, 274: ídem, vista caudal (ec: ectoprocto,
gl: gonapófisis laterales, gs: gonarco, hi: hipandrio, p: pene, pa: parámero, psa: placa subanal, S: esternito, T: terguito).
Escala en mm. Figs. 267-274.— Hemisemidalis pallida, 267: extremo abdominal masculino, vista ventral, 268: ídem, vista lateral (adaptado de Monserrat, 2010), 269-270: variabilidad en el margen caudal del hipandrio y del gonarco, 271: complejo parámeros-pene-décimo esternito, vista
lateral, 272: ídem, vista ventral, 273: extremo abdominal femenino, vista lateral, 274: ídem, vista caudal (ec: ectoprocto,
gl: gonapófisis laterales, gs: gonarco, hi: hipandrio, p: pene, pa: parámero, psa: placa subanal, S: esternito, T: terguito).
Escala en mm.
Figs. 267-274.— Hemisemidalis pallida, 267: male abdominal apex, ventral view, 268: ditto, lateral view (adapted from Monserrat, 2010), 269-270: variability in hypandrium caudal margin and gonarcus, 271: paramera-penis-tenth sternite, lateral view, 272: ditto,
ventral view, 273: female abdominal apex, lateral view, 274: ditto, caudal view (ec: ectoproct, gl: lateral gonapophysis,
gs: gonarcus, hi: hypandrium, p: penis, pa: paramere, psa: subanal plate, S: sternite, T: tergite). Scale in mm.
|
|
|
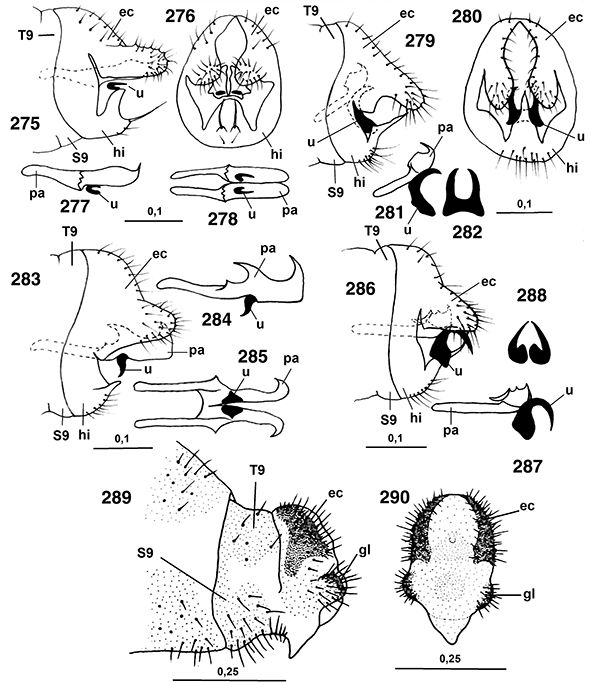 Figs. 275-290.— Genitalia de Semidalis spp. 268-288 (♂). 275-278: S. pluriramosa, 275: extremo abdominal, vista lateral, 276: ídem, vista caudal, 277: parámeros-uncini, vista lateral, 278: ídem, vista ventral,
279-282: S. vicina, 279: extremo abdominal, vista lateral, 280: ídem, vista caudal, 281: parámeros-uncini, vista lateral, 282: uncini, vista
caudal, 283-285: S. aleyrodiformis, 283: extremo abdominal, vista lateral, 284: parámeros-uncini, vista lateral, 285: ídem, vista ventral, 286-288: S. pseudouncinata, 286: extremo abdominal, vista lateral, 287: parámeros-uncini, vista lateral, 288: uncini, vista ventral. 289-290 (♀). 289-290:
extremo abdominal femenino de Semidalis pluriramosa, 289: vista lateral, 290: ídem, vista caudal. Adaptado de Monserrat & Reviejo (1980) (ec: ectoprocto, gl: gonapófisis laterales, hi: hipandrio, pa: parámeros, S: esternito, T: terguito, u: uncini). Escala
en mm. Figs. 275-290.— Genitalia de Semidalis spp. 268-288 (♂). 275-278: S. pluriramosa, 275: extremo abdominal, vista lateral, 276: ídem, vista caudal, 277: parámeros-uncini, vista lateral, 278: ídem, vista ventral,
279-282: S. vicina, 279: extremo abdominal, vista lateral, 280: ídem, vista caudal, 281: parámeros-uncini, vista lateral, 282: uncini, vista
caudal, 283-285: S. aleyrodiformis, 283: extremo abdominal, vista lateral, 284: parámeros-uncini, vista lateral, 285: ídem, vista ventral, 286-288: S. pseudouncinata, 286: extremo abdominal, vista lateral, 287: parámeros-uncini, vista lateral, 288: uncini, vista ventral. 289-290 (♀). 289-290:
extremo abdominal femenino de Semidalis pluriramosa, 289: vista lateral, 290: ídem, vista caudal. Adaptado de Monserrat & Reviejo (1980) (ec: ectoprocto, gl: gonapófisis laterales, hi: hipandrio, pa: parámeros, S: esternito, T: terguito, u: uncini). Escala
en mm.
Figs. 275-290.— Genitalia in Semidalis spp. 268-288 (♂).275-278: S. pluriramosa, 275: abdominal apex, lateral view, 276: ditto, caudal view, 277: paramera-uncini, lateral view, 278: ditto, ventral view,
279-282: S. vicina, 279: abdominal apex, lateral view, 280: ditto, caudal view, 281: paramera-uncini, lateral view, 282: uncini, caudal view,
283-285: S. aleyrodiformis, 283: abdominal apex, lateral view, 284: paramera-uncini, lateral view, 285: ditto, ventral view, 286-288: S. pseudouncinata, 286: abdominal apex, lateral view, 287: paramera-uncini, lateral view, 288: uncini, ventral view. 289-290 (♀). 289-290:
female abdominal apex in Semidalis pluriramosa, 289: lateral view, 290: caudal view. Adapted from Monserrat & Reviejo (1980) (ec: ectoproct, gl: lateral gonapophysis, hi: hypandrium, pa: paramera, S: sternite, T: tergite, u: uncini). Scale in mm.
|
|
|
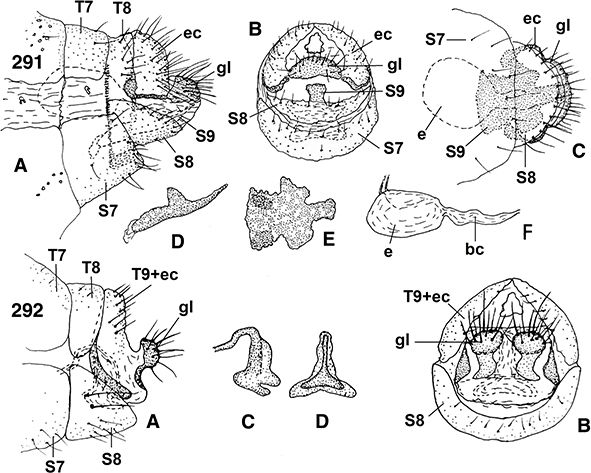 Figs. 291-292.— Estructuras genitales externas e internas en las hembras de Coniopteryx spp.: 291: C. kerzhneri, A: extremo abdominal, vista lateral, B: ídem, vista caudal, C: ídem, vista ventral, D: noveno esternito, vista lateral, E:
ídem, vista ventral, F: espermateca y bolsa copuladora, vista lateral. 292: C. parthenia (sensu Meinander, 1972a), A: extremo abdominal, vista lateral, B: ídem, vista caudal, C: esperpermateca, vista lateral, D: ídem, vista caudal (bc:
bolsa copuladora, e: espermateca, ec: ectoprocto, gl: gonapófisis laterales, S: esternito, T: terguito). A diferentes escalas.
Adaptado de Meinander (1972b). Figs. 291-292.— Estructuras genitales externas e internas en las hembras de Coniopteryx spp.: 291: C. kerzhneri, A: extremo abdominal, vista lateral, B: ídem, vista caudal, C: ídem, vista ventral, D: noveno esternito, vista lateral, E:
ídem, vista ventral, F: espermateca y bolsa copuladora, vista lateral. 292: C. parthenia (sensu Meinander, 1972a), A: extremo abdominal, vista lateral, B: ídem, vista caudal, C: esperpermateca, vista lateral, D: ídem, vista caudal (bc:
bolsa copuladora, e: espermateca, ec: ectoprocto, gl: gonapófisis laterales, S: esternito, T: terguito). A diferentes escalas.
Adaptado de Meinander (1972b).
Figs. 291-292.— External and internal female genitalia in Coniopteryx spp.: 291: C. kerzhneri, A: abdominal apex, lateral view, B: ditto, caudal view, C: ditto, ventral view, D: ninth sternite, lateral view, E: ditto,
ventral view, F: spermatheca and bursa copulatrix, lateral view, 292: C. parthenia (sensu Meinander, 1972a), A: abdominal apex, lateral view, B: ditto, caudal view, C: sperpermatheca, lateral view, D: ditto, caudal view (bc: bursa
copulatrix, e: spermatheca, ec: ectoproct, gl: lateral gonapophysis, S: sternite, T: tergite). At diferent scales. Adapted
from Meinander (1972b).
|
|
|
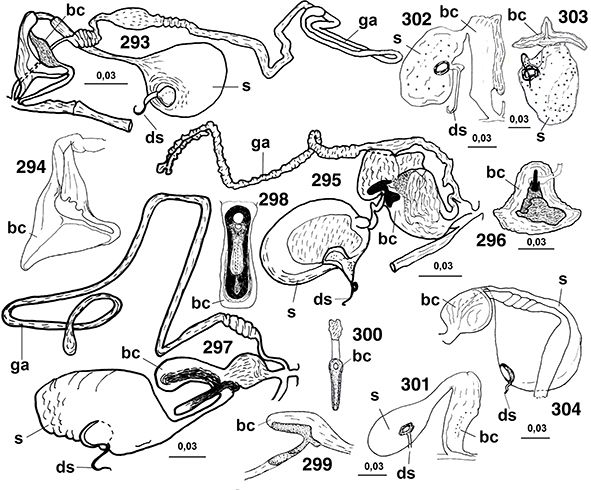 Figs. 293-304.— Estructuras genitales internas en las hembras de Coniopteryx spp.: 293: C. parthenia (sensu Meinander, 1972a), vista lateral, 294: ídem, detalle de la bolsa copuladora, vista lateral, 295: C. drammonti, vista lateral, 296: ídem, detalle de la bolsa copuladora, vista ventral, 297: C. haematica, vista lateral, 298: ídem, detalle de la bolsa copuladora, vista ventral, 299: C. tullgreni, detalle de la bolsa copuladora, vista lateral, 300: ídem, vista ventral, 301: C. tineiformis, vista lateral, 302: C. borealis, vista lateral, 303: ídem, vista ventral, 304: C. pygmaea, vista lateral (bc: bolsa copuladora, ds: ducto seminal, ga: glándula accesoria, s: espermateca). Escalas en mm. Figs. 299,
300: de Tjeder (1964); 293, 295-297, 303: adaptado de Sziráki (1992c); 294, 298, 301, 302, 304: adaptado de Sziráki (1992a). Figs. 293-304.— Estructuras genitales internas en las hembras de Coniopteryx spp.: 293: C. parthenia (sensu Meinander, 1972a), vista lateral, 294: ídem, detalle de la bolsa copuladora, vista lateral, 295: C. drammonti, vista lateral, 296: ídem, detalle de la bolsa copuladora, vista ventral, 297: C. haematica, vista lateral, 298: ídem, detalle de la bolsa copuladora, vista ventral, 299: C. tullgreni, detalle de la bolsa copuladora, vista lateral, 300: ídem, vista ventral, 301: C. tineiformis, vista lateral, 302: C. borealis, vista lateral, 303: ídem, vista ventral, 304: C. pygmaea, vista lateral (bc: bolsa copuladora, ds: ducto seminal, ga: glándula accesoria, s: espermateca). Escalas en mm. Figs. 299,
300: de Tjeder (1964); 293, 295-297, 303: adaptado de Sziráki (1992c); 294, 298, 301, 302, 304: adaptado de Sziráki (1992a).
Figs. 293-304.— Female internal genitalia in Coniopteryx spp.: 293: C. parthenia (sensu Meinander, 1972a), lateral view, 294: ditto, particular of the bursa copulatrix, lateral view, 295: C. drammonti, lateral view, 296: ditto, particular of the bursa copulatrix, ventral view, 297: C. haematica, lateral view, 298: ditto, particular of the bursa copulatrix, ventral view, 299: C. tullgreni, particular of the bursa copulatrix, lateral view, 300: ditto, ventral view, 301: C. tineiformis, lateral view, 302: C. borealis, lateral view, 303: ditto, ventral view, 304: C. pygmaea, lateral view (bc: bursa copulatrix, ds: ductus seminalis, ga: accessory gland, s: spermatheca). Scales in mm. Figs. 299,
300: from Tjeder (1964); 293, 295-297, 303: adapted from Sziráki (1992c); 294, 298, 301, 302, 304: adapted from Sziráki (1992a).
|
|
|
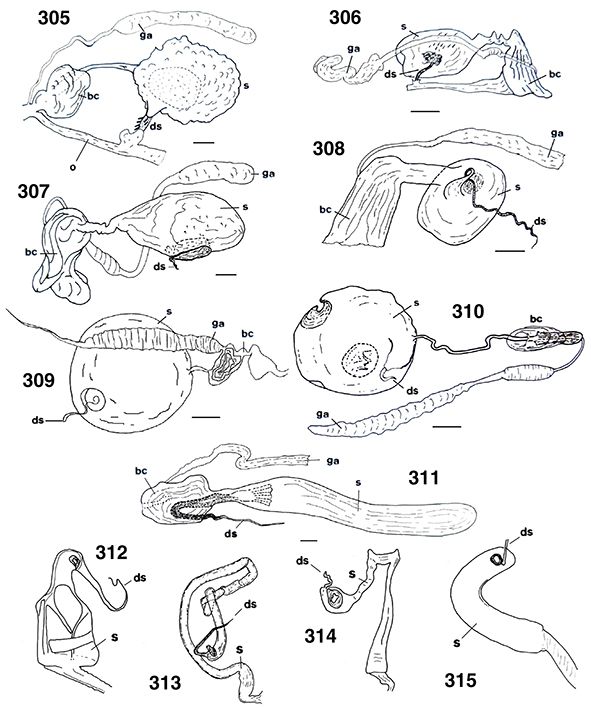 Figs. 305-315.— Estructuras genitales internas en las hembras de: 305: Nimboa adelae, vista lateral, 306: Coniopteryx loipetsederi, vista lateral, 307: Parasemidalis fuscipennis, vista lateral, 308: Hemisemidalis sp., vista lateral, 309: Semidalis pseudouncinata, vista dorsal, 310: Semidalis aleyrodiformis, vista dorsal, 311: Helicoconis lutea, vista lateral, 312: Coniopteryx arcuata, vista ventral, 313: Coniopteryx tjederi, vista lateral, 314: Coniopteryx lentiae, vista ventral, 315: Coniopteryx esbenpeterseni, vista lateral (bc: bolsa copuladora, ds: ducto seminal, ga: glándula accesoria, o: oviducto, s: espermateca). Figs. 305-311:
escala: 0,03 mm; Figs. 305-308, 310-311 adaptado de Sziráki (1996); Figs. 312-315: a diferentes escalas, adaptado y corregido de Sziráki (1992a, 2002a). Figs. 305-315.— Estructuras genitales internas en las hembras de: 305: Nimboa adelae, vista lateral, 306: Coniopteryx loipetsederi, vista lateral, 307: Parasemidalis fuscipennis, vista lateral, 308: Hemisemidalis sp., vista lateral, 309: Semidalis pseudouncinata, vista dorsal, 310: Semidalis aleyrodiformis, vista dorsal, 311: Helicoconis lutea, vista lateral, 312: Coniopteryx arcuata, vista ventral, 313: Coniopteryx tjederi, vista lateral, 314: Coniopteryx lentiae, vista ventral, 315: Coniopteryx esbenpeterseni, vista lateral (bc: bolsa copuladora, ds: ducto seminal, ga: glándula accesoria, o: oviducto, s: espermateca). Figs. 305-311:
escala: 0,03 mm; Figs. 305-308, 310-311 adaptado de Sziráki (1996); Figs. 312-315: a diferentes escalas, adaptado y corregido de Sziráki (1992a, 2002a).
Figs. 305-315.— Female internal genitalia in: 305: Nimboa adelae, lateral view, 306: Coniopteryx loipetsederi, lateral view, 307: Parasemidalis fuscipennis, lateral view, 308: Hemisemidalis sp., lateral view, 309: Semidalis pseudouncinata, dorsal view, 310: Semidalis aleyrodiformis, dorsal view, 311: Helicoconis lutea, lateral view, 312: Coniopteryx arcuata, ventral view, 313: Coniopteryx tjederi, lateral view, 314: Coniopteryx lentiae, ventral view, 315: Coniopteryx esbenpeterseni, lateral view (bc: bursa copulatrix, ds: ductus seminalis, ga: accessory gland, o: oviduct, s: spermatheca). Figs. 305-311:
scale: 0.03 mm; Figs. 305-308, 310-311 adapted from Sziráki (1996); Figs. 312-315: at different scales, adapted and corected from Sziráki (1992a, 2002a).
|
|
AgradecimientosTOP
Esta contribución forma parte y entre 2011-2013 ha contado con fondos del Proyecto coordinado de I+D+i Fauna Ibérica (Neuroptera)
CGL2010-22267-C07-05. Deseamos agradecer a las instituciones, amigos y compañeros citados en el material estudiado por permitirnos
el estudio del material citado de sus colecciones o por el envío de los datos y/o ejemplares de sus colecciones particulares
o de sus centros de investigación, en especial a Fernando Acevedo y Axel Gruppe. También deseamos manifestar nuestro sincero
agradecimiento a las autoridades de las diferentes Comunidades Autónomas por facilitarnos los correspondientes permisos de
muestreos para la toma de datos en los espacios protegidos visitados todos estos años, y en especial a Blanca Ramos del Parque
Nacional y Parque Natural de Sierra Nevada por la cantidad de permisos de muestreos que nos ha facilitado durante todos estos
años. A Vladimir Makarkin por sus valiosas anotaciones, a Roberto A. Pantaleoni y Rinaldo Nicoli Aldini por sus amables intenciones
y comentarios a mis solicitudes, a Hubert y Renate Rausch del Naturkundliche Gesellschaft Mostviertel (Scheibbs, Austria)
y Ben Price del Natural History Museum (Londres, Reino Unido) por ayudarnos a intentar localizar el material original de Janetschek.
A los editores Michel Canard y Henry Pihlström por darnos permiso de reproducción de algunas figuras aquí incluidas, a Isidro
Martínez, Manuel González Núñez y Johann Gepp por sus fotografías, a Fernando Acevedo, Víctor Triviño, Alberto Tinaut, Francisca
Ruano y Rafael Alcalá por su ayuda en los últimos muestreos realizados, a Ángel Rico y David Papenberg por su ayuda en la
traducción de algunos textos, y a Eduardo Ruiz por su apoyo en el tratamiento y composición de las imágenes.
ReferenciasTOP
| ○ |
Ábrahám, L., Markó, V. & Vas, J., 2003. Investigations on a neuropteroid community by using different methods. Acta Phytopathologica et Entomologica Hungarica, 38: 199-207.
|
| ○ |
Ábrahám, L. & Mészáros, Z., 2006. Further Studies on the Daily Activity Pattern of Neuroptera with some Remarks on the Diurnal
Activities. Acta Phytopathologica et Entomologica Hungarica, 41: 275-286.
|
| ○ |
Ábrahám, L. & Vas, J., 1999. Preliminary report on study of the daily activity pattern of Neuroptera in Hungary. Acta Phytopathologica et Entomologica Hungarica, 34: 153-164.
|
| ○ |
Acker, T. S., 1960. The comparative morphology of the male terminalia of Neuroptera (Insecta). Microentomology, 24: 25-83.
|
| ○ |
Adams, P. A., 1973. Coniopterygidae from Chile, with Three New Species (Neuroptera). The Pan-Pacific Entomologist, 49(3): 250-254.
|
| ○ |
Agekyan, N. G., 1978. A little known entomophagous insect Semidalis aleyrodiformis Stephens (Neuroptera, Coniopterygidae) in Adzharia. Entomologicheskoe Obozrenie, 57: 509-512,
|
| ○ |
Alayo, D. P., 1968. Los Neurópteros de Cuba. Poeyana. Serie B, 2: 1-127.
|
| ○ |
Albarda, H., 1889. Catalogue raisonné et synonymique des Nevroptères, observés dans les Pays-Bas et dans les Pays limitrophes.
Tijdschrift voor Entomologie, 32: 211-376.
|
| ○ |
Alvis, L., Villalba, M., Marzal, C. & García-Marí, F., 2003. Identification and abundance of Neuropteran species associated
with citrus orchards in Valencia, Spain. Integrated Control in Citrus Fruit Crops, 26(6): 185-190.
|
| ○ |
Andersen, T. & Greve, L., 1975. Neuroptera in light-traps at Osterøy, Hordaland. Norwegian Journal of Entomology, 22: 123-128.
|
| ○ |
Anónimo, 1918 (probablemente L. Navás). Insectos nuevos de Cataluña. Boletín de la Sociedad entomológica de España, 1(8): 172.
|
| ○ |
Ansorge, J., 1996. Insekten aus dem Oberen Lias von Grimmen (Vorpommern, Norddeutschland). Neue Palaontologische Abhandlungen, 2: 1-132.
|
| ○ |
Arrow, G. J., 1917. The life-history of Conwentzia psociformis Curt. The Entomologist’s Monthly Magazine, 53: 254-257.
|
| ○ |
Askew, R. R., 1971. Parasitic insects. American Elsevier. New York. 316 pp.
|
| ○ |
Aspöck, H., 1963a. Coniopteryx loipetsederi nov. spec. (Neuroptera, Coniopterygidae). Nachrichtenblatt der Bayerischen Entomologen, 12: 95.
|
| ○ |
Aspöck, H., 1963b. Coniopteryx tjederi Kimmins-ein für Mitteleuropa neues Neuropteron (Ein Beitrag zur Kenntnis der Coniopterygiden Österreichs). Nachrichtenblatt der Bayerischen Entomologen, 12: 41-44.
|
| ○ |
Aspöck, H., 1964. Coniopteryx hölzeli nov. spec., ein neues europäisches Neuropteron. Entomologische Berichten, 24: 77-78.
|
| ○ |
Aspöck, H., 1992. The Neuropteroidea of Europe: a review of present knowledge (Insecta). In: Canard, M., Aspöck, H. & Mansell,
M. W. (eds.). Current Research in Neuropterology (Proceedings of the Fourth International Symposium on Neuropterology, Bagnères-de-Luchon
1991). Toulouse: 43-56.
|
| ○ |
Aspöck, H. & Aspöck, U., 1964. Synopsis der Systematik, Ökologie und Biogeographie der Neuropteren Mitteleuropas im Spiegel
der Neuropteren-Fauna von Linz und Oberösterreich, sowie Bestimmungsschlüssel für die mitteleuropäischen Neuropteren und Beschreibung
von Coniopteryx lentiae nov. spec. Naturkundliches Jahrbuch der Stadt Linz, 1964: 127-282.
|
| ○ |
Aspöck, H. & Aspöck, U., 1965a. Coniopteryx pinkeri nov. spec. von den Kanarischen Inseln (Mit Bemerkungen über eine homogene Arten-Gruppe des Genus Coniopteryx Curtis). Zeitschrift der Arbeitsgemeinschaft Österreichischer Entomologen, 17: 79-85.
|
| ○ |
Aspöck, H. & Aspöck, U., 1965b. Vorläufige Mitteilung über die Coniopterygiden Vorderasiens (Neuroptera). Entomologisches Nachrichtenblatt, 12: 17-23.
|
| ○ |
Aspöck, H. & Aspöck, U., 1969. Die Neuropteren Mitteleuropas: ein Nachtrag zur “Synopsis der Systematik, Ökologie und Biogeographie
der Neuropteren Mitteleuropas.” Naturkundliches Jahrbuch der Stadt Linz, 1969: 17-68.
|
| ○ |
Aspöck, H. & Aspöck, U., 2003. Early History of Coniopterygid Research. Abstracts of the 8th International Symposium on Neuropterology, Texas: 19-20.
|
| ○ |
Aspöck, H. & Aspöck, U., 2014. Coniopteryx (C.) pygmaea Enderlein, 1906, and Coniopteryx (C.) hoelzeli H. Aspöck, 1964 – a final statement (Insecta: Endopterygota: Neuropterida: Neuroptera). DGaaE-Nachrichten, 28(1): 6-9.
|
| ○ |
Aspöck, H., Aspöck, U. & Hölzel, H., 1980. Die Neuropteren Europas, 2 vols. Goecke & Evers. Krefeld, t. I, 495 pp, t. II, 355 pp.
|
| ○ |
Aspöck, H. & Hölzel, H., 1996. The Neuropteroidea of North Africa, Mediterranean Asia and of Europe: a comparative review
(Insecta). In: Canard, M., Aspöck, H. & Mansell, M. W. (eds.). Pure and Applied Research in Neuropterology (Proceedings of the Fifth International Symposium on Neuropterology, Cairo 1994). Toulouse: 31-86.
|
| ○ |
Aspöck, H., Hölzel, H. & Aspöck, U., 2001. Kommentierter Katalog der Neuropterida (Insecta: Raphidioptera, Megaloptera, Neuroptera)
der Westpaläarktis. Denisia, 2: 1-606.
|
| ○ |
Aspöck, U., 1992. Crucial points in the phylogeny of the Neuroptera (Insecta). In: Canard, M., Aspöck, H. & Mansell, M. W.
(eds.). Current Research in Neuropterology (Proceedings of the Fourth International Symposium on Neuropterology, Bagnères-de-Luchon
1991). Toulouse: 63-73.
|
| ○ |
Aspöck, U., 1995. Neue hypothesen zum System der Neuropterida. Mitteilungen der Deutschen Gesellschaft für Allgemeine und Angewandte Entomologie, 10: 633-636.
|
| ○ |
Aspöck, U., 2002. Phylogeny of the Neuropterida (Insecta: Holometabola). Zoologica Scripta, 31(1): 51-55. http://dx.doi.org/10.1046/j.0300-3256.2001.00087.x |
| ○ |
Aspöck, U., 2003. Kamelhälse, Schlammfliegen, Netzflügler--zur Phylogenie der Neuropterida (Insecta: Holometabola). Verhandlungen Westdeutscher Entomologentag, 14: 7-9.
|
| ○ |
Aspöck, U. & Aspöck, H., 1999. Kamelhälse, Schlammfliegen, Ameisenlöwen. Wer sind sie? (Insecta: Neuropterida: Raphidioptera,
Megaloptera, Neuroptera). Stapfia, 60: 1-34.
|
| ○ |
Aspöck, U. & Aspöck, H., 2007. Verbliebene Vielfalt vergangener Blüte. Zur Evolution, Phylogenie und Biodiversität der Neuropterida
(Insecta: Endopterygota). Denisia, 20: 451-516.
|
| ○ |
Aspöck, U. & Aspöck, H., 2008. Phylogenetic relevance of the genital sclerites of Neuropterida (Insecta: Holometabola). Systematic Entomology, 33: 97-127. http://dx.doi.org/10.1111/j.1365-3113.2007.00396.x |
| ○ |
Aspöck, U. & Aspöck, H., 2010. Landmarks towards a phylogeny-based classification of the Neuropterida (Insecta: Endopterygota).
In: Devetak, D., Lipovšek, S. & Arnett, A. E. (eds.). Proceedings of the 10th International Symposium on Neuropterology, Piran 2008. University of Maribor. Maribor: 67-74.
|
| ○ |
Aspöck, U. & Aspöck, H., 2013. Nevrorthidae, Sisyridae, Coniopterygidae. Der Streit um die niederen Ränge im Stammbaum der Neuroptera. DGaaE Nachrichten, 27: 19-20.
|
| ○ |
Aspöck, U., Aspöck, H. & Haring, E., 2003. Phylogeny of the Neuropterida-morphological evidence and the molecular advocatus
diaboli. Entomologische Abhandlungen, 61: 157-158.
|
| ○ |
Aspöck, U., Aspöck, H., Letardi, A. & Jong, Y de, 2015. Fauna Europaea: Neuropterida (Raphidioptera, Megaloptera, Neuroptera).
Biodiversity Data Journal, 3: e4830. http://dx.doi.org/10.3897/BDJ.3.e4830 |
| ○ |
Aspöck, U., Haring, E. & Aspöck, H., 2012. The phylogeny of the Neuropterida: long lasting and current controversies and challenges
(Insecta: Endopterygota). Arthropod Systematics & Phylogeny, 70: 119-129.
|
| ○ |
Aspöck, U. & Mansell, M. W., 1994. A revision of the family Rhachiberothidae Tjeder, 1959, stat. n. (Neuroptera). Systematic Entomology, 19: 181-206. http://dx.doi.org/10.1111/j.1365-3113.1994.tb00587.x |
| ○ |
Aspöck, U., Plant, J. D. & Nemeschkal, H. L., 2001. Cladistic analysis of Neuroptera and their systematic position within
Neuropterida (Insecta: Holometabola: Neuropterida: Neuroptera). Systematic Entomology, 26: 73-86. http://dx.doi.org/10.1046/j.1365-3113.2001.00136.x |
| ○ |
Azar, D., Nel, A. [D.] & Solignac, M., 2000. A new Coniopterygidae from the Lebanese amber. Acta Geológica Hispanica, 35: 31-36.
|
| ○ |
Badgley, M. E., Fleschner, C. A. & Hall, J. C., 1955. The biology of Spiloconis picticornis Banks (Neuroptera: Coniopterygidae). Psyche, 62: 75-81. http://dx.doi.org/10.1155/1955/16126 |
| ○ |
Báez, M. & Oromí, P., 2010. Planipennia. In: Arechavaleta Hernández, M. et al. (Coords.). Lista de especies silvestres de Canarias (hongos, plantas y animales terrestres). Gobierno de Canarias. La Laguna: 253-254.
|
| ○ |
Bagnall, R. S., 1915a. A brief review of the British Coniopterygidae (Neuroptera), with tables of the European genera and
species. Entomologist’s Record and Journal of Variation, 27: 241-247.
|
| ○ |
Bagnall, R. S., 1915b. Conwentzia cryptoneuris sp. n., a neuropteron (Coniopterygidae) new to the British fauna. The Entomologist’s Monthly Magazine, 51: 192-193.
|
| ○ |
Balduf, W. V., 1939. The bionomics of entomophagous Insects. Vol. 2. John S. Swift Co. Chicago. [iii] + 384 pp.
|
| ○ |
Banks, N., 1892. A synopsis, catalogue, and bibliography of the neuropteroid insects of temperate North America. Transactions of the American Entomological Society, 19: 327-373.
|
| ○ |
Banks, N., 1899. Descriptions of new North American neuropteroid insects. Transactions of the American Entomological Society, 25: 199-218.
|
| ○ |
Banks, N., 1906. A revision of the Nearctic Coniopterygidae. Proceedings of the Entomological Society of Washington, 8: 77-86.
|
| ○ |
Banks, N., 1913. Synopses and descriptions of exotic Neuroptera. Transactions of the American Entomological Society, 39: 201-242.
|
| ○ |
Barnard, P. C., Brooks, S. J. & Stork, N. E., 1986. The seasonality and distribution of Neuroptera, Raphidioptera and Mecoptera
on oaks in Richmond Park, Surrey, as revealed by insecticide knock-down sampling. Journal of Natural History, 20: 1321-1331. http://dx.doi.org/10.1080/00222938600770881 |
| ○ |
Bastero Monserrat, J. J., 1989. Longinos Navás, científico jesuita. Universidad de Zaragoza. Zaragoza. 229 pp.
|
| ○ |
Beché, J. R., 1913. Excursión a Pina (Zaragoza). Boletín de la Sociedad Aragonesa de Ciencias Naturales, 12: 167-170.
|
| ○ |
Berland, L., 1962. Atlas des Névroptères de France, Belgique, Suisse. Mégaloptères, Raphidioptères, Névroptères Planipennes, Mécoptères, Trichoptères. Nérée Boubée & Cie. Paris. 158 pp.
|
| ○ |
Beutel, R. G., Friedrich, F. & Aspöck, U., 2010. The larval head of Nevrorthidae and the phylogeny of Neuroptera (Insecta).
Zoological Journal of the Linnean Society, 158: 533-562. http://dx.doi.org/10.1111/j.1096-3642.2009.00560.x |
| ○ |
Billberg, G. J., 1820. Enumeratio Insectorum in Museo Gust. Joh. Billberg. Typis Gadelianis. Holmiae. 138 pp.
|
| ○ |
Blasco, J., 2000. Le Genévrier thurifère (Juniperus thurifera L.) dans le bassin occidental de la Méditerranée: systématique, écologie, dynamique et gestion. Les Dossiers forestiers, 6: 94-103.
|
| ○ |
Bodenheimer, F. S., 1929. Materialien zur Geschichte der Entomologie bis Linné. Vol. 2. W. Junk. Berlin. 486 pp.
|
| ○ |
Bohigas, M. & Sanchez, A., 1917. Catáleg dels insectes del Museu pertanyents als orders: Paraneuroptera-Ephemeroptera-Plecoptera-Neuroptera-Megaloptera-Rhaphidioptera-Embioptera-Mecoptera-Psocoptera-Trichoptera.
Publicacions de la Junta de Ciències Naturals de Barcelona, Anuari, 2(1): 301-326.
|
| ○ |
Brauer, F., 1852. Versuch einer Gruppirung der Gattungen in der Zunft Planipennia mit besonderer Rücksicht auf die früheren
Stände. Stettiner Entomologische Zeitung, 13: 71-77.
|
| ○ |
Brauer, F., 1855a. Beiträge zur Kenntniss des inneren Baues und der Verwandlung der Neuropteren. Verhandlungen des Zoologisch-Botanischen Vereins in Wien, 5: 701-726.
|
| ○ |
Brauer, F., 1855b. Beiträge zur Kenntniss der Verwandlung der Neuropteren. Verhandlungen des Zoologisch-Botanischen Vereins in Wien, 5: 777-786.
|
| ○ |
Brauer, F., 1868. Verzeichniss der bis jetzt bekannten Neuropteren in Sinne Linné’s. Erster Abschnitt. Verhandlungen der Kaiserlich-Königlichen Zoologisch-Botanischen Gesellschaft in Wien, 18: 359-416, 711-742.
|
| ○ |
Brauer, F., 1876. Die Neuropteren Europas und insbesondere Oesterreichs mit Rücksicht auf ihre geographische Verbreitung.
Festschrift zur Feier des fünfundzwanzigjährigen Bestehen Kaiserlich-Koniglichen Zoologisch-Botanischen Gesellschaft: 263-300.
|
| ○ |
Brauer, F., 1900. Über die von Prof. O. Simony auf den Canaren gefundenen Neuroptera und Pseudoneuroptera (Odonata, Corrodentia
et Ephemeridae). Sitzungsberichte der Akademie der Wissenschaften in Wien, Mathematische-Naturwissenschaftliche Klasse (Abtheilung I), 109: 464-477.
|
| ○ |
Brauer, F. & Löw, F., 1857. Neuroptera austriaca. Die im Erzherzogthum Oesterreich bis jetzt aufgefundenen Neuropteren nach der analytischen Methode zusammengestellt,
nebst einer kurzen Charakteristik aller europäischen Neuropteren-Gattungen. C. Gerold’s Sohn. Wien. xxiii + 80 pp.
|
| ○ |
Britten, H., 1916. Semidalis (Coniopteryx) aleurodiformis Steph., on the wing. The Entomologist’s Monthly Magazine, 52: 43.
|
| ○ |
Burmeister, H. C. C., 1839. Handbuch der Entomologie. Zweiter Band. Besondere Entomologie. Zweite Abtheilung. Kaukerfe. Gymnognatha. Zweite Hälfte; vulgo Neuroptera. Theodor C.F. Enslin. Berlin. xii + 757-1050 pp.
|
| ○ |
Canard, M., 2001. Natural food and feeding habits of lacewings. In: McEwen, P. K., New, T. R. & Whittington, A. E. (eds.).
Lacewings in the Crop Environment. Cambridge University Press. Cambridge: 116-129.
|
| ○ |
Canard, M. & Volkovich, T. A., 2001. Outlines of lacewing development. In: McEwen, P. K., New, T. R. & Whittington, A. E.
(eds.). Lacewings in the Crop Environment. Cambridge University Press. Cambridge: 130-153.
|
| ○ |
Canbulat, S., 2007. Contributions to the knowledge of the lacewing fauna of Turkey (Neuroptera: Coniopterygidae, Myrmeleontidae),
with some ecological remarks. Entomological News, 118(3): 303-310. http://dx.doi.org/10.3157/0013-872X(2007)118[303:CTTKOT]2.0.CO;2 |
| ○ |
Canbulat, S., 2013. Two new records of the Coniopteryx (Neuroptera: Coniopterygidae) species for the Turkish fauna. Turkish Journal of Zoology, 37: 242-245. http://dx.doi.oeg/10.3906/zoo-1205-10 |
| ○ |
Carpenter, F. & Lestage, J. A., 1928. Une sous-famille nouvelle (Fontenelleinae) du groupe des Coniopterygoidea Till. Recueil de l’Institut Zoologique Torley-Rousseau, 1: 153-172.
|
| ○ |
Carpenter, F. M., 1930. A Review of our present knowledge of the geological history of the insects. Psyche, 37(1): 15-34. http://dx.doi.org/10.1155/1930/37616 |
| ○ |
Carpenter, F. M., 1955. Redescription of Spiloconis picticornis Banks (Neuroptera, Coniopterygidae). Psyche, 62(2): 69-74. http://dx.doi.org/10.1155/1955/58729 |
| ○ |
Carpenter, F. M., 1992. Treatise on Invertebrate Paleontology. Part R. Arthropoda 4. Vols. 3 and 4: Superclass Hexapoda. The Geological Society of America and The University of Kansas. xxi + 277 pp. [vol 3], ii + 279-655 [vol 4].
|
| ○ |
Castellari, P. L., 1980. Indagini biologiche su Coniopteryx (Metaconiopteryx) esbenpeterseni Tjeder (Neur. Coniopterygidae), predatore di Acari Tetranichidi sul Pesco. Bollettino dell’Istituto di Entomologia della Università degli Studi di Bologna, 35:157-180.
|
| ○ |
Cheng, L. & Birch, M. C., 1978. Insects flotsam: an unstudied marine resource. Ecological Entomology, 3: 87-97. http://dx.doi.org/10.1111/j.1365-2311.1978.tb00907.x |
| ○ |
Collyer, E., 1951. The separation of Conwentzia pineticola End. from Conwentzia psociformis (Curt.), and notes on their biology. Bulletin of Entomological Research, 42: 555-564. http://dx.doi.org/10.1017/S0007485300028959 |
| ○ |
Collyer, E., 1964. Phytophagous mites and their predators in New Zealand ochards. New Zealand Journal of Agricultural Research, 7(4): 551-568.
|
| ○ |
Crampton, G. C., 1921. The sclerites of the head, and the mouthparts of certain immature and adult insects. Annals of the Entomological Society of America, 14: 65-110. http://dx.doi.org/10.1093/aesa/14.2.65 |
| ○ |
Curtis, J., 1834. British entomology; being illustrations and descriptions of the genera of insects found in Great Britain and Ireland: containing
coloured figures from nature of the most rare and beautiful species, and in many instances of the plants upon which they are
found. Vol. 11. Lovell Reeve & Co. London. 769 lám.
|
| ○ |
Cuvier, G. L. C. F. D., 1817. Le règne animal distribué d’après son organisation, pour servir de base à l’histoire naturelle des animaux et d’introduction
a l’anatomie comparée. 4 Vols. Deterville. Paris.
|
| ○ |
Czechowska, W., 1982. Neuroptera and Mecoptera of Warsaw and Mazovia. Memorabilia Zoologica, 36: 165-184.
|
| ○ |
Czechowska, W., 1986. Structure of Neuropteran (Neuropteroidea) communities in urban green areas of Warsaw. Memorabilia Zoologica, 41: 187-214.
|
| ○ |
De Geer, C., 1778. Mémoires pour servir à l’histoire des insectes. Tome VII. Hosselberg. Stockholm. 950 pp.
|
| ○ |
De Geer, C., 1783. Genera et species insectorum e generossimi auctoria scriptis. Lebrecht. Lipsiae. 220 pp.
|
| ○ |
Dessart, P., 1978. À propos de deux espèces de Dendrocerus (Atritomellus) (Hym. Ceraphronoidea Megaspilidae) parasites de Coniopterix esbenpeterseni (Neuroptera) en Italie. Bulletin et Annales de la Société Royale Entomologique de Belgique, 114:177-182.
|
| ○ |
Dessart, P., 1998. Notules hyménoptérologiques n°s 22-26. Bulletin et Annales de la Société Royale Entomologique de Belgique, 133: 403-418.
|
| ○ |
Devetak, D., 1992. Rdeči seznam ogroženih mrežekrilcev (Neuroptera s. l.) v Sloveniji. Varstvo Narave, 17: 111-115.
|
| ○ |
Devetak, D., 1998. Detection of substrate vibration in Neuropteroidea: a review. In: Panelius, S. P. (ed.). Neuropterology 1997 (Proceedings of the Sixth International Symposium on Neuropterology, Helsinki 1997). Acta Zoologica Fennica, 209: 87-94.
|
| ○ |
Díaz-Aranda, L. M. & Monserrat, V. J., 1988a. Contribución al conocimiento de los neurópteros de Jaén (Insecta, Neuroptera).
Boletín de la Asociación Española de Entomología, 12: 111-123.
|
| ○ |
Díaz-Aranda, L. M. & Monserrat, V. J., 1988b. Contribución al conocimiento de los neurópteros de Teruel (Insecta, Neuropteroidea).
Boletín de la Asociación Española de Entomología, 12: 215-231.
|
| ○ |
Díaz-Aranda, L. M. & Monserrat, V. J., 1988c. Contribución al conocimiento de los Neurópteros de Granada (Insecta, Neuropteroiea).
Actas del III Congreso Ibérico de Entomología, Granada: 211-227.
|
| ○ |
Díaz-Aranda, L.M., Monserrat, V. J. & Marín, F., 1986a. Composición biogeográfica de los neurópteros de Guadalajara (Insecta,
Neuropteroidea: Planipennia). Actas de las VIII Jornadas de la Asociación Española de Entomología, Sevilla: 1121-1130.
|
| ○ |
Díaz-Aranda, L.M., Monserrat, V. J. & Marín, F., 1986b. Contribución al conocimiento de los neurópteros de Guadalajara (Insecta,
Neuropteroidea). Actas de las VIII Jornadas de la Asociación Española de Entomología, Sevilla: 1131-1144.
|
| ○ |
Dinkins, R. L., Tedders, W. L. & Reid, W., 1994. Predaceous neuropterans in Georgia and Kansas Pecan Trees. Journal of Entomological Science, 29: 165-175.
|
| ○ |
Dobosz, R. & Krzeminski, W., 2000. A new species of the Coniopterygidae (Neuroptera) from Baltic amber. Polskie Pismo Entomologiczne, 69: 219-224.
|
| ○ |
Domingo-Quero, T., Alonso-Zarazaga, M. A., Sánchez-Ruiz, A., Araujo, R., Navas, A., Sánchez Moreno, S., García Becerra, R.,
Nebreda, M., Sánchez Ruiz, M., Fontal-Cazalla, F., Nieves-Aldrey, J.L., 2003. Inventariando la biodiversidad en el Parque
Nacional de La Caldera de Taburiente (La Palma, Islas Canarias, España): novedades científicas. Graellsia, 59(2-3): 45-68. http://dx.doi.org/10.3989/graellsia.2003.v59.i2-3.235 |
| ○ |
Dorokhova, G. I., 1987. [Order Neuroptera.] Opredeliteli po faune SSSR, 153: 36-96. (en ruso).
|
| ○ |
Duelli, P., 1994a. Rote Liste der gefährdeten Netzflügler der Schweiz. In: Duelli, P. Rote Listen der gefährdeten Tierarten der Schweiz. Bundesamt für Umwelt, Wald und Landschaft, BUWAL-Reihe Rote Listen und EDMZ. Bern: 64-65.
|
| ○ |
Duelli, P., 1994b. Neuroptera from the Mediterranean Island of Malta (Neuroptera). Entomofauna, 15: 125-127.
|
| ○ |
Duelli, P., Obrist, M. K. & Flückiger, P. F., 2002. Forest edges are biodiversity hotspots--also for Neuroptera. In: Sziráki,
G. (ed.). Neuropterology 2000 (Proceedings of the Seventh International Symposium on Neuropterology, Budapest 2000). Acta Zoologica Academiae Scientiarum Hungaricae, 48 (Suppl. 2): 75-87.
|
| ○ |
Dujardin, F., 1851. Sur une larve qui paraît être celle de l’Hemerobius hirtus. Annales des Sciences Naturelles, Zoologie et Biologie Animale, (3), 15: 169-173.
|
| ○ |
Dziędzielewicz, J., 1905. Bielotki Galicyi i S’laska. Kosmos (Lwów), 30: 377-385.
|
| ○ |
Dziędzielewicz, J., 1907. Przyczynek do systematyki Bielotek (Coniopterygidae). Kosmos (Lwów), 32: 208-214.
|
| ○ |
Eglin, W., 1940. Die Neuropteren der Umgebung von Basel. Revue Suisse de Zoologie, 47: 243-358.
|
| ○ |
Eglin, W., 1982a. Zur Insektenfauna der Umgebung der Vogelwarte Sempach, Kanton Luzern. III. Neuropteroidea und Mecoptera
(Netzflügler und Schnabelfliegen). Entomologische Berichte Luzern, 7: 62-70.
|
| ○ |
Eglin, W., 1982b. Zur Insektenfauna der Umgebung des Brisen-Haldigrates, 1200-2400 m, und vom Pilatus-Kulm, 2060 m, Kanton
Nidwalden. V. Neuropteroidea und Mecoptera (Netzflügler und Schnabelfliegen). Entomologische Berichte Luzern, 8: 57-60.
|
| ○ |
Emerton, J. H., 1906. Cocoons and young of Coniopteryx vicina. Psyche, 13: 74-75. http://dx.doi.org/10.1155/1906/624860 |
| ○ |
Enderlein, G., 1905a. Klassifikation der Neuropteren-familie Coniopterygidae. Zoologischer Anzeiger, 29: 225-227.
|
| ○ |
Enderlein, G., 1905b. Conwentzia pineticola nov. gen. nov. spec. eine neue Neuroptere aus Westpreussen. Bericht des Westpreussischen Botanischen-Zoologischen Vereins, 26/27(Anlagen): 10-12.
|
| ○ |
Enderlein, G., 1905c. Ein neuer zu den Coniopterygiden gehöriger Neuropteren-Typus aus der Umgebung von Berlin. Wiener Entomologische Zeitung, 24: 197-198.
|
| ○ |
Enderlein, G., 1906. Monographie der Coniopterygiden. Zoologische Jahrbücher. Abteilung für Systematik, Geographie und Biologie, 23: 173-242.
|
| ○ |
Enderlein, G., 1907a. Die Coniopterygidenfauna Japans. Stettiner Entomologische Zeitung, 68: 3-9.
|
| ○ |
Enderlein, G., 1907b. Eine neue Coniopterygide aus Java. Notes from the Leyden Museum, 28: 224-226.
|
| ○ |
Enderlein, G., 1907c. Nachträge zur monographie der Coniopterygiden. Stettiner Entomologische Zeitung, 68: 10-13.
|
| ○ |
Enderlein, G., 1908. Neuroptera Fam. Coniopterygidae. Genera Insectorum, 67: 1-18.
|
| ○ |
Enderlein, G., 1909. Zur Kenntnis frühjurassischer Copeognathen und Coniopterygiden und über das Schicksal der Archipsylliden.
Zoologischer Anzeiger, 34: 770-776.
|
| ○ |
Enderlein, G., 1910a. Über de Beziehungen der fossilen Coniopterygiden zu den recenten und über Archiconiocompsa prisca nov. gen. nov. spec. Zoologischer Anzeiger, 35: 673-677.
|
| ○ |
Enderlein, G., 1910b. Embiidina und Neuroptera (Coniopterygidae und Hemerobiidae). Transactions of the Linnean Society of London, Zoology, (2), 14(1): 55-58. http://dx.doi.org/10.1111/j.1096-3642.1910.tb00523.x |
| ○ |
Enderlein, G., 1914. Ueber zwei neue afrikanische Coniopterygiden. Bollettino del Laboratorio di Entomologia Agraria di Portici, 8: 225-227.
|
| ○ |
Enderlein, G., 1929. Entomologica Canaria II. Zoologischer Anzeiger, 84: 221-234.
|
| ○ |
Enderlein, G., 1930. Die Klassifikation der Coniopterygiden auf Grund der recenten und fossilen Gattungen. Archiv für Klassifikatorische und Phylogenetische Entomologie, 1: 98-114.
|
| ○ |
Engel, M. S., 2002. A new dustywing (Neuroptera: Coniopterygidae) in Turonian amber from New Jersey, with a reassessment of
Glaesoconis in Neocomian amber from Lebanon. Journal of the Kansas Entomological Society, 75: 38-42.
|
| ○ |
Engel, M. S., 2004. The dustywings in Cretaceous Burmese amber (Insecta: Neuroptera: Coniopterygidae). Journal of Systematic Palaeontology, 2: 133-136, 98 (plate). http://dx.doi.org/10.1017/S1477201904001191 |
| ○ |
Engel, M. S., 2010. A new genus of dustywings allied to Archiconiocompsa in Baltic amber (Neuroptera: Coniopterygidae). Transactions of the Kansas Academy of Sciences, 113: 145-150.
|
| ○ |
Engel, M. S. & Grimaldi, D. A., 2007. The neuropterid fauna of Dominican and Mexican amber (Neuropterida, Megaloptera, Neuroptera).
American Museum Novitates, 3587: 1-58. http://dx.doi.org/10.1206/0003-0082(2007)3587
|
| ○ |
Engel, M. S. & Grimaldi, D. A., 2008. Diverse Neuropterida in Cretaceous amber, with particular reference to the paleofauna
of Myanmar (Insecta). Nova Supplementa Entomologica, 20: 1-86.
|
| ○ |
Esben-Petersen, P., 1918. Results of Dr. E. Mjöberg’s Swedish Scientific Expeditions to Australia 1910-1913. 18. Neuroptera
and Mecoptera. Arkiv för Zoologi, 11(26): 1-37.
|
| ○ |
Esben-Petersen, P., 1928. Neuroptera and Embiidina from Abyssinia and Somaliland. Annals and Magazine of Natural History, (10), 1: 442-450. http://dx.doi.org/10.1080/00222932808672808 |
| ○ |
Esben-Petersen, P., 1931. Inventa entomologica itineris Hispanici et Maroccani, quod a. 1926 fecerunt Harald et Håkan Lindberg.
VIII. Neuroptera. Commentationes Biologicae, 3(13): 1-6.
|
| ○ |
Fabricius, J. C., 1775. Systema entomologiae, sistens insectorvm classes, ordines, genera, species, adiectis synonymis, locis, descriptionibvs, observationibvs. Offic. Libr. Kortii. Flensbvrgi et Lipsiae. 832 pp.
|
| ○ |
Fabricius, J. C., 1777. Genera insectorvm eorvmqve characteres natvrales secvndvm nvmervm, figvram, sitvm et proportionem omnivm partivm oris adiecta
mantissa speciervm nvper detectarvm. M. F. Bartsch. Chilonii. 310 pp.
|
| ○ |
Fabricius, J. C., 1778. Philosophia entomologica sistens scientiae fvndamenta adiectis definitionibvs, exemplis, observationibvs, advmbrationibvs. C. E. Bohn. Hambvrgi ei Kilonii. [xii] + 178 pp.
|
| ○ |
Fabricius, J. C., 1781. Species insectorvm exhibentes eorvm differentias specificas, synonyma avctorvm, loca natalia, metamorphosin adiectis observationibvs,
descriptionibvs. Tome 1. C. E. Bohnii. Hamburgi et Kilonii. 552 pp.
|
| ○ |
Fabricius, J. C., 1793. Entomologia systematica emendata et aucta secundum classes, ordines, genera, species adjectis synonimis, locis observationibus,
descriptionibus. Tome 2. C. G. Proft. Hafniae. 519 pp.
|
| ○ |
Fabricius, J. C., 1798. Supplementum entomologiae systematicae. Proft et Storch. Hafniae. 572 pp.
|
| ○ |
Fitch, A., 1854. Report [upon the noxious and other insects of the state of New-York]. Transactions of the New York State Agricultural Society, 14: 705-880.
|
| ○ |
Fitch, A., 1856. First and second report on the noxious, beneficial and other insects, of the state of New-York. C. van Benthuysen, New York. [ii] + 336 pp.
|
| ○ |
Fleschner, C. A., 1950. Studies on searching capacity of the larvae of three predators of the citrus red mite. Hilgardia, 20: 233-265.
|
| ○ |
Fleschner, C. A. & Ricker, D. W., 1953. Food habits of Coniopterygids on citrus in southern California. Journal of Economic Entomology, 46: 458-461. http://dx.doi.org/10.1093/jee/46.3.458 |
| ○ |
Fraser, F.C., 1952. New additions to the fauna of Madagascar. Odonata and Neuroptera. Mémoires de l’Institut Scientifique de Madagascar. Série E. Entomologie, 1: 135-143.
|
| ○ |
Fraser, F.C., 1955. Nouvelles notes sur les Nevropteres de Madagascar. Naturaliste Malgache, 7: 127-137.
|
| ○ |
Fraser, F.C., 1957. Odonata and Neuroptera of Reunion. Mémoires de l’Institut Scientifique de Madagascar. Série E. Entomologie, 8: 15-28.
|
| ○ |
Fraser, F.C., 1959. Mecoptera, Megaloptera and Neuroptera. Handbooks for the identification of British Insects, 1(12-13): 1-40.
|
| ○ |
Frazer, B. D., 1988. Predators. In: Minks, A.K. & Harrewijn, P. (eds). Aphids, their biology, natural enemies and control, Vol. 2B. Amsterdam. Elsevier: 217-230.
|
| ○ |
García Marí, F. & González Zamora, J.E., 1999. Biological control of Tetranychus urticae (Acari: Tetranychidae) with naturally occurring predators in strawberry plantings in Valencia, Spain. Experimental and Applied Acarology, 23: 487-495. http://dx.doi.org/10.1023/A:1006191519560 |
| ○ |
García Marí F., Santaballa, E., Ferragut, F., Marzal, C., Colomer, P. & Costa, J., 1983. El ácaro rojo Panonychus citri (McGregor): incidencia en la problemática fitosanitaria de nuestros agrios. Boletín de Sanidad Vegetal, Plagas, 9: 191-218.
|
| ○ |
Genay, A., 1953. Contribution à l’étude des Névroptères de Bourgogne. Travaux du Laboratoire de Zoologie et de la Station Aquicole Grimaldi de la Faculté des Sciences de Dijon (N.S.), 3: 1-30.
|
| ○ |
Gepp, J., 1967. Die Coniopterygidae des Grazer Feldes und seiner Randgebiete (Neuroptera). Mitteilungen des Naturwissenschaftlichen Vereins für Steiermark, 97: 76-80.
|
| ○ |
Gepp, J., 1969. Eine neue Coniopterygidenart aus Anatolien: Aleuropteryx perpusilla n. sp. (Neuroptera, Planipennia). Zeitschrift der Arbeitsgemeinschaft Österreichischer Entomologen, 21: 12-15.
|
| ○ |
Gepp, J., 1974a. Semidalis vicina in Südosteuropa (Planipennia, Coniopterygidae). Entomologische Zeitschrift, 84: 223-225.
|
| ○ |
Gepp, J., 1974b. Coniopterygiden aus Jugoslawien (Planipennia). Zeitschrift der Arbeitsgemeinschaft Österreichischer Entomologen, 24: 167-168.
|
| ○ |
Gepp, J., 1977. Die Planipennier der Steiermark (Neuroptera s. str., Neuropteroidea, Insecta): Autökologie und Regionalfaunistik.
Mitteilungen des Naturwissenschaftlichen Vereins für Steiermark, 107: 171-206.
|
| ○ |
Gepp, J., 1981. Rote Liste der gefährdeten Netzflüglerarten der Steiermark (Megaloptera, Raphidioptera und Planipennia). Steirischer Naturschutzbrief, Sonderheft, 3: 91-96.
|
| ○ |
Gepp, J., 1983. Rote Liste der gefährdeten Netzflügler Österreichs (Megaloptera, Raphidioptera und Planipennia). In: Rote Listen gefährdeter Tiere Österreichs. Bundesministeriums für Gesundheit und Umweltschutz. Wien: 145-147.
|
| ○ |
Gepp, J., 1984. Erforschungsstand der Neuropteren-Larven der Erde (mit einem Schlüssel zur larvaldiagnose der Familien, einer
Übersicht von 340 beschreibenen Larven und 600 Literaturzitaten). In: Gepp J, Aspöck H, Hölzel H (Eds.). Progress in World’s Neuropterology (Proceedings of the 1st International Symposium on Neuropterology, Graz 1980). Graz: 183-239.
|
| ○ |
Gepp, J., 1986. Biology and larval diagnosis of central European Neuroptera (a review of present knowledge). In: Gepp, J.,
Aspöck, H. & Hölzel, H. (Eds.). Recent Research in Neuropterology (Proceedings of the 2nd International Symposium on Neuropterology, Hamburg 1984. Graz: 137-144.
|
| ○ |
Gepp, J., 1990. An illustrated review of egg morphology in the families of Neuroptera (Insecta: Neuropteroidea). In: Mansell,
M. W. & Aspöck, H. (eds.). Advances in Neuropterology (Proceedings of the Third International Symposium on Neuropterology, Berg en Dal 1988). South African Department of Agricultural Development. Pretoria: 131-149.
|
| ○ |
Gepp, J., 1992. Contribution to the knowledge of coniopterygid larvae of Central Europe (Insecta: Neuroptera: Coniopterygidae).
In: Canard, M., Aspöck, H. & Mansell, M. W. (eds.). Current Research in Neuropterology (Proceedings of the Fourth International Symposium on Neuropterology, Bagnères-de-Luchon
1991). Toulouse: 151-152.
|
| ○ |
Gepp, J., 1999. Neuropteren als Indikatoren der Naturraumbewertung. Eignung als Modellgruppe, Methodenwahl, Fallbeispiele
sowie Diskussion möglicher Fragestellungen (Neuropterida). Stapfia, 60: 167-208.
|
| ○ |
Gepp, J., Hellrigl, K. & Hölzel, H., 1994. Lista rossa dei neurotteri (Neuropteroidea) minacciati dell’Alto Adige. In: Lista Rossa delle specie animali minacciate in Alto Adige. Anonymous. Provincia Autonoma di Bolzano/Alto Adige: 302-307.
|
| ○ |
Gepp, J. & Stürzer, C., 1986. Semidalis aleyrodiformis (Steph., 1836) – Biologie, Ökologie und Larvenstadien (Planipennia, Coniopterygidae). Mitteilungen des Naturwissenschaftlichen Vereins für Steiermark, 116: 241-262.
|
| ○ |
Gerling, D., Alomar, Ò. & Arnó, J., 2001. Biological control of Bemisia tabaci using predators and parasitoids. Crop Protection, 20: 779-799. http://dx.doi.org/10.1016/S0261-2194(01)00111-9 |
| ○ |
Ghilarov, M. S., 1962. The larva of Dilar turcicus Hag. and the position of the family Dilaridae in the order Planipennia. Entomologicheskoe Obozrenie, 41: 402-416.
|
| ○ |
Ghosh, S. K. & Sen, S., 1977. Check-list of Indian Planipennia (Order Neuroptera). Records of the Zoological Survey of India, 73: 277-326.
|
| ○ |
González Olazo, E. V., 1984a. Stangesemidalis subandina, nuevo genero y especie de Coniopterygidae de la Republica Argentina (Neuroptera, Planipennia). Acta Zoologica Lilloana, 38: 59-63.
|
| ○ |
González Olazo, E. V., 1984b. Contribucion al conocimiento de Semidalis absurdiceps (Enderlein, 1908), (Neuroptera, Coniopterygidae).
Acta Zoologica Lilloana, 37: 187-189.
|
| ○ |
González Olazo, E. V., 1987. Una nueva especie de Coniopteryx (Coniopterygidae, Neuroptera) del Brasil. Acta Zoologica Lilloana, 39: 11-12.
|
| ○ |
Greve, L., 1974. The larvae and pupa of Helicoconis lutea (Wallengren, 1871) (Neuroptera, Coniopterygidae). Norsk Entomologisk Tidsskrift, 21: 19-23.
|
| ○ |
Greve, L. & Kobro, S., 1998. Preliminary data on abundance of phototactic Neuroptera and Rephidioptera in SE Norway as indicated
by light-trap catches. In: Panelius, S. P. (ed.). Neuropterology 1997 (Proceedings of the Sixth International Symposium on Neuropterology, Helsinki 1997). Acta Zoologica Fennica, 209: 119-120.
|
| ○ |
Grimaldi, D. A., 2000. A diverse fauna of Neuropterodea in amber from the Cretaceous of New Jersey. In: Grimaldi, D. (ed.).
Studies on fossils in amber, with particular reference to the Cretaceous of New Jersey. Backhuys Publishers. Leiden: 259-303.
|
| ○ |
Grimaldi, D. A. & Engel, M. S., 2005. Evolution of the Insects. Cambridge University Press. New York. xv + 755 pp.
|
| ○ |
Grimaldi, D. A., Engel, M. S. & Nascimbene, P. C., 2002. Fossiliferous Cretaceous amber from Myanmar (Burma): its rediscovery,
biotic diversity, and peleontological significance. American Museum Novitates, 3361: 1-72. http://dx.doi.org/10.1206/0003-0082(2002)361%3C0001:FCAFMB%3E2.0.CO;2 |
| ○ |
Grimaldi, D., Engel, M.S., Nascimbene, P.C. & Singh, H., 2013. Coniopterygidae (Neuroptera: Aleuropteryginae) in amber from
the Eocene of India and the Miocene of Hispaniola. American Museum Novitates, 3770: 1-20. http://dx.doi.org/10.1206/3770.2 |
| ○ |
Gruppe, A., 2007a. Spatial distribution of Neuropterida in the LAK stand: significance of host tree specifity. In: Unterseher,
M., Morawetz, W., Klotz, S. & Arndt, E. (eds.). The canopy of a temperate floodplain forest. Results from five years of research at the Leipzig Canopy Crane. Department of Systematic Botany, Botanical Garden, and Herbarium Universtitatis Lipsiensis. Leipzig: 91-96.
|
| ○ |
Gruppe, A., 2007b. Ungewöhnliche Funde von Neuropteren. DGaaE-Nachrichten, 21(3): 141.
|
| ○ |
Günther, K. K., 1991. Echte Netzflügler aus Lichtfängen im Stadtgebiet von Berlin (Insecta, Neuroptera). Entomologische Nachrichten und Berichte, 35: 161-170.
|
| ○ |
Günther, K. K., 1993. Welche Art muss Coniopteryx pygmaea Enderlein, 1906 heissen? (Neuroptera, Coniopterygidae). Deutsche Entomologische Zeitschrift (N.F.), 40: 167-171.
|
| ○ |
Hagen, H. A., 1851. Uebersicht der neueren Literatur, betreffend die Neuroptera Linn. Stettiner Entomologische Zeitung, 12:118-128, 185-191, 345-350, 365-375.
|
| ○ |
Hagen, H. A., 1852. Uebersicht der neueren Literatur, betreffend die Neuroptera Linn. Stettiner Entomologische Zeitung, 13: 90-95, 108-115, 154-158, 230-233.
|
| ○ |
Hagen, H. A., 1854. Ueber die Neuropteren der Bernsteinfauna. Verhandlungen des Zoologisch-Botanischen Vereins in Wien, 4: 221-232.
|
| ○ |
Hagen, H. A., 1856. Die im Bernstein befindlichen Neuropteren der vorwelt bearbeitet von F. J. Pictet-Baraban, und Dr. H.
Hagen. In: Berendt, G. C. (ed). Die im Bernstein befindlichen organischen reste der Vorwelt gesammelt, in verbindung mit mehreren bearbeitet. Vol. 2. Nicholaischen Buchhandlung. Berlin: 41-125.
|
| ○ |
Hagen, H. A., 1858a. Synopsis of the British Planipennes. Entomologist’s Annual, 1858: 17-33.
|
| ○ |
Hagen, H. A., 1858b. Synopsis der Neuroptera Ceylons. Verhandlungen der Kaiserlich-Königlichen Zoologisch-Botanischen Gesellschaft in Wien, 8: 471-488.
|
| ○ |
Hagen, H. A., 1859a. Synopsis der Neuroptera Ceylons (Pars II.). Verhandlungen der Kaiserlich-Königlichen Zoologisch-Botanischen Gesellschaft in Wien, 9: 199-212.
|
| ○ |
Hagen, H. A., 1859b. On Coniopteryx. Entomologist’s Weekly Intelligencer, 5: 189.
|
| ○ |
Hagen, H. A., 1861. Synopsis of the Neuroptera of North America, with a list of the South American species. Smithsonian Miscellaneous Collections, 4(1): xx + 1-347. http://dx.doi.org/10.5962/bhl.title.60275 |
| ○ |
Hagen, H. A., 1866. Hemerobidarum Synopsis Synonymica. Stettiner Entomologische Zeitung, 27: 369-462.
|
| ○ |
Hagen, H. A., 1873. On the larvae of the Hemerobina. Proceedings of the Boston Society of Natural History, 15: 243-248.
|
| ○ |
Handlirsch, A., 1906. Revision of American Paleozoic insects. Proceedings of the United States National Museum, 29: 661-820. http://dx.doi.org/10.5479/si.00963801.29-1441.661 |
| ○ |
Handlirsch, A., 1906-1908. Die fossilen Insekten und die Phylogenie der rezenten Formen. W. Engelmann, Leipzig. 1430 pp. (Para años de publicación de sus apartados puede consultarse Oswald, 2013a, 2013b).
|
| ○ |
Handlirsch, A., 1909. Zur Kenntnis “frühjurassischer Copeognathen und Coniopterygiden” und über das Schiksal der Archipsylliden.
Zoologischer Anzeiger, 35: 233-240.
|
| ○ |
Haring, E. & Aspöck, U., 2004. Phylogeny of the Neuropterida: a first molecular approach. Systematic Entomology, 29: 415-430. http://dx.doi.org/10.1111/j.0307-6970.2004.00263.x |
| ○ |
Haring, E., Aspöck, H., Bartel, D. & Aspöck, U., 2011. Molecular phylogeny of the Raphidiidae (Raphidioptera). Systematic Entomology, 36(1): 16-30. http://dx.doi.org/10.1111/j.1365-3113.2010.00542.x |
| ○ |
Hennig, W., 1969. Phylogenetic Systematics. University of Illinois Press. Urbana. 263 pp.
|
| ○ |
Henry, C. S., 1982. Neuroptera. In: Parker, S. (ed.). Synopsis and Classification of Living Organisms. Vol. 2. McGraw-Hill. New York: 470-482.
|
| ○ |
Henry, T. J., 1976. Aleuropteryx juniperi: a European scale predator established in North America (Neuroptera: Coniopterygidae). Proceedings of the Entomological Society of Washington, 78: 195-201.
|
| ○ |
Hinton, H. E, 1981. Megaloptera and Neuroptera. In: Biology of insect eggs. Vol. 2. Pergamon Press. Oxford: 642-643.
|
| ○ |
Hoffmann, J., 1962. Faune des Névroptéroïdes du Grand-Duché de Luxembourg. Archives, Institut Grand-Ducal de Luxembourg, Section des Sciences Naturelles, Physiques et Mathématiques (N.S.), 28: 249-332.
|
| ○ |
Hollier, J. A. & Belshaw, R. D., 1993. Stratification and phenology of a woodland Neuroptera assemblage. The Entomologist, 112: 169-175.
|
| ○ |
Hölzel, H., 1964. Die Netzflügler Kärntens. Carinthia II, 74: 97-156.
|
| ○ |
Hölzel, H., 1973. Die Netzflügler Kärntens. 1. Nachtrag. Carinthia II, 83: 497-506.
|
| ○ |
Hölzel, H., 1998. Zoogeographical features of Neuroptera of the Arabian peninsula. In: Panelius, S. P. (ed.). Neuropterology 1997 (Proceedings of the Sixth International Symposium on Neuropterology, Helsinki 1997). Acta Zoologica Fennica, 209: 129-140.
|
| ○ |
Hölzel, H., 1999. Verzeichnis der Netzfluegler Kaerntens In: Rottenburg, T., Wieser, C., Mildner, P. & Holzinger, W. E. (eds.).
Rote Listen gefaehrdeter Tiere Kaerntens. Band 15. Naturschutz. Klagenfurt: 413-416.
|
| ○ |
Horn, W., Kahle, I., Friese, G. & Gaedike, R., 1990. Collectiones entomologicae. Ein Kompendium über den Verbleib entomologischer Sammlungen der Welt bis 1960. Akademie der Lanwirtschaftswissenschaften der Deutschen Demokratischen Republik. Berlin. 2 Vols.: vol. 1: 220 pp.; vol. 2:
221-573 pp.
|
| ○ |
Houser, J. S., 1914. Conwentzia hageni Banks. Life history notes and variations in wing venation. Annals of the Entomological Society of America, 7: 73-76. http://dx.doi.org/10.1093/aesa/7.1.73 |
| ○ |
Hynd, W. R. B., 1989. Coniopteryx lentiae Aspöck & Aspöck (Neuroptera: Coniopterygidae) new to Britain. Entomologist’s Gazette, 40: 159-160.
|
| ○ |
ICZN (International Commission on Zoological Nomenclature). Opinion 1595, 1990. Aleuropteryx Loew, 1885 (Inseca, Neuroptera): Aleuropteryx loewii Klapalak, 1894 designated as the type species. Bulletin of Zoological Nomenclature, 47(2): 153.
|
| ○ |
Ivanov, V. D. & Krivokhatsky, V. A., 1999. Insects and spiders of the Leningrad Region. In: Balashova, N.B. & Zavarzin, A.A.
Biodiversity of the Leningrad Region (Algae. Fungi. Lichens. Bryophytes. Invertebrates. Fisches and pisciformes. St. Petersburg University Press. St. Petersburg: 339-354.
|
| ○ |
Ivanov, V. D. & Krivokhatsky, V. A., 2002. Insects. In: Noskov, G. A. (ed.). Red data book of nature of the Leningrad region. Biological Research Institute of the St. Petersburg State University and World & Family. Saint Petersburg: 95-300.
|
| ○ |
Ivanov, V. D. & Krivokhatsky, V. A., 2004. Insecta. In: Noskov, G. A. (ed.). Red data book of nature of Saint-Petersburg. Biological Research Institute of the St. Petersburg State University. Saint-Petersburg: 179-242.
|
| ○ |
Izquierdo Moya, I., Martín Albaladejo, C., Díaz de Castro, J. & González Galán, A., 2010. Catálogo de los tipos de especies
de hexápodos descritas de las islas Canarias conservados en el Museo Nacional de Ciencias Naturales (Madrid, España). Vieraea, 38: 23-54.
|
| ○ |
Janetschek, H., 1957. Zoologische Ergebnisse einer Studienreise in die spanische Sierra Nevada. Publicaciones del Instituto de Biología Aplicada, 26: 135-153.
|
| ○ |
Jedlička, L., 2001. Red (Ecosozological) List of Neuroptera of Slovakia. In: Baláž, D., Marhold, K. & Urban, P. (eds.). Red List of Plants and Animals of Slovakia. Ochrana Prírody. Banská Bystrica: 108-109.
|
| ○ |
Jepson, J. E. & Penney, D., 2007. Neuropteran (Insecta) palaeodiversity with predictions for the Cretaceous fauna of the Wealden.
Palaeogeography, Palaeoclimatology, Palaeoecology, 248: 109-118. http://dx.doi.org/10.1016/j.palaeo.2006.11.015 |
| ○ |
Johnson, V., 1976. A new genus and species of Coniopterygidae (Neuroptera) from New Mexico. Psyche, 83: 192-195. http://dx.doi.org/10.1155/1976/57125 |
| ○ |
Johnson, V., 1977. A review of the Coniopterygidae of North and Central America. Ph.D. dissertation. University of Kentucky.
Lexington. xii + 185 pp.
|
| ○ |
Johnson, V., 1978. New Coniopterygidae from Central America. Annals of the Entomological Society of America, 71: 163-164. http://dx.doi.org/10.1093/aesa/71.2.163 |
| ○ |
Johnson, V., 1980a. Review of the Coniopterygidae (Neuroptera) of North America with a revision of the genus Aleuropteryx. Psyche, 87: 259-298. http://dx.doi.org/10.1155/1980/62163 |
| ○ |
Johnson, V., 1980b. Notes on Coniopterygidae (Neuroptera) from the Ethiopian region. Entomological News, 91: 159-160.
|
| ○ |
Johnson, V., 1981. New species of Coniopterygidae (Neuroptera) from North America. Psyche, 87:181-192. http://dx.doi.org/10.1155/1980/42021 |
| ○ |
Johnson, V. & Morrison, W. P., 1979. Mating behavior of three species of Coniopterygidae (Neuroptera). Psyche, 86: 395-398. http://dx.doi.org/10.1155/1979/27503 |
| ○ |
Joost, W., 1973. Neuropteren aus Bulgarien. Entomologische Nachrichten, 17: 145-156.
|
| ○ |
Karny, H. H., 1923. Eine neue Coniopteryx aus Buitenzorg (Neur. Megalopt.). Treubia, 3: 384-386.
|
| ○ |
Karny, H. H., 1924. On a remarkable new coniopterygid genus from Egypt (Neuroptera Megaloptera). Annals and Magazine of Natural History, (9), 13: 474-478. http://dx.doi.org/10.1080/00222932408633060 |
| ○ |
Killington, F. J., 1929. A synopsis of British Neuroptera. Transactions of the Entomological Society of Hampshire and the South of England, 5: 1-36.
|
| ○ |
Killington, F. J., 1932. The parasites of Neuroptera with special reference to those attacking British species. Transactions of the Entomological Society of the South of England, 8: 84-91.
|
| ○ |
Killington, F.J., 1936. A monograph of the British Neuroptera I. Royal Society. London. 269 pp.
|
| ○ |
Killington, F.J., 1937a. A monograph of the British Neuroptera II. Royal Society. London. 306 pp.
|
| ○ |
Killington, F. J., 1937b. The generic names of the British Neuroptera with a check list of the British species. Generic Names of British Insects, 4: 66-80.
|
| ○ |
Kimmins, D. E., 1934. A new species of Coniopteryx (Neuroptera) from France. Annals and Magazine of Natural History, (10), 13: 613-619. http://dx.doi.org/10.1080/00222933408654863 |
| ○ |
Kimmins, D. E., 1935. Some new South African Neuroptera. Annals and Magazine of Natural History, (10), 15: 561-579. http://dx.doi.org/10.1080/00222933508655001 |
| ○ |
Kimmins, D. E., 1950. A brachypterous coniopterygid (Order Neuroptera) from Mt. Ruwenzori, Uganda. Annals and Magazine of Natural History, (12), 3: 166-171. http://dx.doi.org/10.1080/00222935008654049 |
| ○ |
Kimmins, D. E., 1952. New species of East African Neuroptera. The Entomologist, 85: 186-190.
|
| ○ |
Kimmins, D. E., 1953. New Hawaiian Coniopterygidae (Neuroptera). Proceedings of the Hawaiian Entomological Society, 15: 37-39.
|
| ○ |
Kimmins, D. E., 1960. On a small collection of Neuroptera from Glorioso Island, with a description of a new species of Coniopterycidae
[sic]. Naturaliste Malgache, 12: 113-115.
|
| ○ |
Kimmins, D. E. & Wise, K. A. J., 1962. A record of Chryptoscenea australiensis (Enderlein) (Neuroptera: Coniopterygidae) in New Zealand, with a re-description of the species. Transactions of the Royal Society of New Zealand, Zoology, 2: 35-39.
|
| ○ |
Kis, B., 1964a. Eine neue Aleuropteryx-Art aus Rumänien (Neuroptera, Coniopterygidae). Reichenbachia, 4: 227-230.
|
| ○ |
Kis, B., 1964b. Zur Kenntnis der Coniopterygiden Rumäniens (Neuroptera, Coniopterygidae). Entomologische Abhandlungen, 31: 49-56.
|
| ○ |
Kis, B., 1965a. Die Helicoconis-Arten Rumäniens (Neuroptera, Coniopterygidae). Reichenbachia, 5: 125-130.
|
| ○ |
Kis, B., 1965b. Eine neue Coniopteryx-Art aus Rumänien (Neuroptera, Coniopterygidae). Reichenbachia, 5: 295-298.
|
| ○ |
Kis, B., 1965c. Beiträge zur Kenntnis der Neuropterenfauna Rumäniens. Rovartani Közlemények (N.S.), 18: 359-366.
|
| ○ |
Kis, B., 1967. Coniopteryx aspöcki n. sp., eine neue Neuropterenart aus Europa. Reichenbachia, 8: 123-125.
|
| ○ |
Kis, B., 1969. Genul Coniopteryx Curt. (Neuroptera) în România. Comunicari de Zoologie, 2: 35-41.
|
| ○ |
Kis, B., 1972. Ord. Neuroptera (l’entomofaune du “Grind” de Caraorman, delta du Danube). Travaux du Muséum d’Histoire Naturelle “Grigore Antipa”, 12: 125-130.
|
| ○ |
Kis, B., 1975. Neuroptera. In: Ionescu, M. (ed.). Fauna. Seria Monografică. Academiei Republicii Socialiste România, Porţiele de Fier. Bucarest: 98-100.
|
| ○ |
Kis, B., Nagler, C. & Mândru, C., 1970. Insecta: Neuroptera (Planipennia). Fauna Republicii Socialiste România, 8(6): 1-343.
|
| ○ |
Klapálek, F., 1894. Is Aleuropteryx lutea, Löw, identical with Coniopteryx lutea Wallg.? The Entomologist’s Monthly Magazine, 30: 121-122.
|
| ○ |
Klapálek, F., 1909. Některé zajímavější Neuropteroidy španělské. Acta Societatis Entomologicae Bohemiae, 6: 130-131.
|
| ○ |
Kleinsteuber, E., 1974. Verzeichnis der im Gebiet der Deutschen Demokratischen Republik bisher festgestellten Neuropteren
(Neuropteroidea: Megaloptera, Raphidioptera et Planipennia). Entomologische Nachrichten, 18: 145-153.
|
| ○ |
Klingstedt, H., 1936. Die Coniopterygiden der Kanarischen Inseln. Commentationes Biologicae, 6(8): 1-5.
|
| ○ |
Krüger, L., 1915. Neuroptera. Eine historisch-systematische übersicht. I. Neuroptera L. bis Neuroptera Erichs. Stettiner Entomologische Zeitung, 76: 3-51.
|
| ○ |
Krüger, L., 1917. Neuroptera. Eine historische-systematische übersicht. II. Phylogenetische Umgrenzung der Echten Neuropteren:
Neuropteroidea Hdl. Stettiner Entomologische Zeitung, 78: 116-137.
|
| ○ |
Krüger, L., 1923. Neuroptera succinica baltica. Die im baltischen Bernstein eingeschlossenen Neuropteren des Westpreussischen
Provinzial-Museums (heute Museum für Naturkunde und Vorgeschichte) in Danzig. Stettiner Entomologische Zeitung, 84: 68-92.
|
| ○ |
Kubrakiewicz, J., Jedrzejowska, I. & Bilinski, S. M., 1998. Neuropteroidea – different ovary structure in related groups.
Folia Histochemica et Cytobiologica, 36: 179-187.
|
| ○ |
Kupryjanowicz, J. & Makarkin, V. N., 2008. Archiconiocompsa prisca Enderlein (Neuroptera: Coniopterygidae): the first neuropteran fossil in Rovno amber (Ukraine). Entomologica Fennica, 19: 25-31.
|
| ○ |
Kuwayama, S., 1962. A revisional synopsis of the Neuroptera in Japan. Pacific Insects, 4(2): 325-412.
|
| ○ |
Lackschewitz, P., 1929. Nachtrag zu den Neuropteren und Trichopteren des Ostbaltischen Gebietes [Mecoptera Comst., Megaloptera
Latr., Planipennia Latr., Trichoptera Kirby.]. Eesti Loodusteaduse Arhiiv, (2), 14(4): 1-40.
|
| ○ |
Lacroix, J. L., 1924. Faune des Planipennes de France. Coniopterygidae. Bulletin de la Société d’Étude des Sciences Naturelles d’Elbeuf, 42: 53-84.
|
| ○ |
Laidlaw, F. F., 1923. The Neuroptera Planipennia recorded from Somerset: with some recent records from Devon and Gloucester.
Proceedings of the Somersetshire Archaeological and Natural History Society, 69: 54-69.
|
| ○ |
Larsson, S. G., 1978. Baltic Amber-a palaeobiological Study. Entomonograph, 1: 1-192.
|
| ○ |
Latreille, P. A., 1810. Considérations générales sur l’ordre naturel des animaux composant les classes des Crustacés, des Arachnides, et des Insectes;
avec un tableau méthodique de leurs genres, disposés en familles. Schoell. Paris. 444 pp.
|
| ○ |
Lauterbach, K. E., 1970. Die Planipennier oder echten Netzflügler der Umgebung von Tübingen. Veröffentlichungen der Landesstelle für Naturschutz und Landschaftspflege Baden-Württemberg, 38: 113-133.
|
| ○ |
Lauterbach, K. E., 1972. Die Planipennier oder echten Netzflügler der Umgebung von Tübingen (Insecta-Neuroptera). Veröffentlichungen der Landesstelle für Naturschutz und Landschaftspflege Baden-Württemberg, 40: 141-144.
|
| ○ |
Leach, W. E., 1815. Entomology. Neuroptera. In: Brewster, D. (ed.). Edinburgh Encyclopaedia, 9(1): 136-139.
|
| ○ |
Letardi, A., 2002a. A web page on Italian Neuropterida. Acta Zoologica Academiae Scientiarum Hungaricae, 48 (Suppl. 2): 155-158.
|
| ○ |
Letardi, A., 2002b. Banche dati in rete: l´esempio dei Neurotteri (Insecta Neuropterida). Atti XIX Congresso nazionale italiano di Entomologia, Catania: 225-229.
|
| ○ |
Letardi, A., 2010. Sharing entomological knowledge: eight years after the release of the Italian Neuropterological web site. In: Devetak, D., Lipovšek, S. & Arnett, A. E. (eds.). Proceedings of the 10th International Symposium on Neuropterology, Piran 2008. University of Maribor. Maribor: 175-179.
|
| ○ |
Letardi, A., 2012. Dilar pumilus Navás, 1903 and Conwentzia pineticola (Enderlein, 1905), two new lacewings for Continental Portugal (Neuroptera: Dilaridae, Coniopterygidae). Arquivos Entomolóxicos, 7: 71-72.
|
| ○ |
Letardi, A., Almeida, J. M., Badano, D., Silva, R. R. & Machado, E., 2013. Contributing to a Checklist of Neuropterida in
Portugal: The Naturdata Project. Açoreana, 2013(Suplemento 9): 29-38.
|
| ○ |
Lewis, T. & Taylor, L. R., 1964. Diurnal periodicity of flight by insects. Transactions of the Royal Entomological Society of London, 116: 393-469. http://dx.doi.org/10.1111/j.1365-2311.1965.tb02304.x |
| ○ |
Linnaeus, C., 1758. Systema natura per regna tria naturae secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis,
locis. Editio decima, reformata. Tomus I. Salvii. Holmiae. 824 pp.
|
| ○ |
Liu, Z.-q., 1995. Two new record genera and new species of Coniopteryginae (Neuroptera: Coniopterygidae) from China. Entomotaxonomia, 17(Suppl.): 35-38.
|
| ○ |
Liu, Z.-q., 2003. Studies on the taxonomy and the taxonomic information system of Coniopterygidae from China. Ph.D. dissertation. China Agricultural University. Beijing. iii + 226 pp.
|
| ○ |
Liu, Z.-q. & Yang, C.-k., 1993. Four new species of Coniopteryginae (Neuroptera: Coniopterygidae) from Guizhou province. Entomotaxonomia, 15: 255-260.
|
| ○ |
Liu, Z.-q. & Yang, C.-k., 1997. Neuroptera: Coniopterygidae. In: Yang, X.-k. (ed.). Insects of the Three Gorge Reservoir area of Yangtze river. Vol. 1. Chongqing Publishing House. Chongqing: 575-579.
|
| ○ |
Liu, Z.-q. & Yang, C.-k., 1998. New species and new records of Coniopterygidae from north China (Neuroptera). Acta Entomologica Sinica, 41(Supp.): 186-193.
|
| ○ |
Liu, Z.-q. & Yang, C.-k., 2001. Neuroptera: Coniopterygidae and Berothidae. In: Wu, H. & Pan, C.-w. (eds.). Insects of Tianmushan National Nature Reserve. Science Press. Beijing: 295-296.
|
| ○ |
Liu, Z.-q. & Yang, C.-k., 2002. Neuroptera: Coniopterygidae. In: Huang, F.-s. (chief ed.). Forest insects of Hainan. Science Press. Beijing: 302-304.
|
| ○ |
Liu, Z. & Yang, J., 2006. Coniopterygidae. In: Li, Z.-z. & Jin, D.-c. (eds.). Insects from Fanjingshan landscape. Guizhou Science and Technology Publishing House. Guiyang: 439-440.
|
| ○ |
Liu, Z.-q., Yang, C.-k. & Shen, Z.-r., 2003a. The first record of genus Aleuropteryx Löw (Neuroptera: Coniopterygidae) from China with description of a new species. Entomotaxonomia, 25: 197-200.
|
| ○ |
Liu, Z.-q., Yang, C.-k. & Shen, Z.-r., 2003b. Two new species of Coniopterygidae (Neuroptera) from Yunnan, China. Entomotaxonomia, 25: 143-147.
|
| ○ |
Liu, Z.-q., Yang, C.-k. & Shen, Z.-r., 2004a. A study of the genus Heteroconis Enderlein (Insecta: Neuroptera: Coniopterygidae) with four new species from mainland China. Raffles Bulletin of Zoology, 52: 365-372.
|
| ○ |
Liu, Z.-q., Yang, C.-k. & Shen, Z.-r., 2004b. A review of the genus Coniocompsa Enderlein (Neuroptera: Coniopterygidae) from China, with descriptions of three new species. Oriental Insects, 38: 395-404.
|
| ○ |
Lo Verde, G. & Monserrat, V. J., 1997. Nuovi dati sui Coniopterygidae Siciliani (Neuropteroidea Planipennia). Il Naturalista Siciliano, (4), 21: 57-66.
|
| ○ |
Lock, K., 2015. Coniopteryx (Metaconiopteryx) esbenpeterseni Tjeder, 1930 new to Belgium (Neuroptera: Coniopterygidae). Bulletin de la Société royale belge d’Entomologie, 151: 115-117.
|
| ○ |
Löw, F., 1885. Beitrag zur Kenntniss der Coniopterygiden. Sitzungsberichte der Akademie der Wissenschaften in Wien, Mathematische-Naturwissenschaftliche Klasse (Abtheilung I), 91: 73-89.
|
| ○ |
Macho Bariego, V., 1909. Excursión anual de la Sociedad Aragonesa de Ciencias Naturales al Pirineo Aragonés (8-15 Julio de
1907). Boletín de la Sociedad Aragonesa de Ciencias Naturales, 8: 67-79.
|
| ○ |
Macleod E. G., 1964. A comparative morphological study of the head capsule and cervix of larval Neuroptera (Insecta). Ph.D. dissertation. Harvard University. Cambridge. [iii] + 528 pp. 35 lám.
|
| ○ |
Makarkin, V. N., 1990. A check-list of the Neuroptera-Planipennia of the USSR far east, with some taxonomic remarks. Acta Zoologica Hungarica, 36: 37-45.
|
| ○ |
Mani, M. S., 1968. Ecology and Biogeography of High Altitude Insects. Series entomologica vol 4. W. Junk. The Hague. XVI + 528 pp.
|
| ○ |
Mansell, M.W., 1986. 19. Order Neuroptera. In: C. H. Scholtz & E. Holm (eds.). Insects of Southern Africa. Butterworths. Durban: 181-187.
|
| ○ |
Mansell, M. W., 2010. Towards a catalogue of Afrotropical Lacewings and Alderflies (Neuroptera, Megaloptera). In: Devetak, D., Lipovšek, S. & Arnett, A. E. (eds.). Proceedings of the 10th International Symposium on Neuropterology, Piran 2008. University of Maribor. Maribor: 1-10.
|
| ○ |
Marcet, A., 1948-1953. Flora Montserratina. Boletín de la Real Sociedad Española de Historia Natural, 46: 331-410, 523-581; 47: 67-89, 163-170, 305-333, 469-484,625-651; 48: 87-108, 327-338; 49: 201-264; 50: 299-379; 51: 51-109.
|
| ○ |
Marín, F., 1994. Las comunidades de neurópteros de la provincia de Albacete (Insecta: Neuropteroidea). Al-Basit, Estudios albacetenses, 34: 247-304.
|
| ○ |
Marín, F. & Monserrat, V. J., 1987. Los neurópteros del encinar ibérico (Insecta, Neuropteroidea). Boletín de Sanidad Vegetal, Plagas, 13: 347-359.
|
| ○ |
Marín, F. & Monserrat, V. J., 1989. Contribución al conocimiento de los neurópteros de Cantabria (Insecta, Neuropteroidea).
Boletín de la Asociación Española de Entomología, 13: 287-298.
|
| ○ |
Marín, F. & Monserrat, V. J., 1990. Contribución al conocimiento de los Neurópteros de Soria (Insecta, Neuropteroidea). Boletín Asociación española Entomología, 14: 219-230.
|
| ○ |
Marín, F. & Monserrat, V. J., 1991a. The Community of Neuropteroidea from Iberian southern beechwoods. In: L. Polgar, R.J.
Chambers, A.F.G. Dixon & I. Hodek (eds). Behaviour and impact of Aphidophaga (Proceedings of the 4th meeting of the IOBC). SPB Academic Publishing. The Hague: 187-198.
|
| ○ |
Marín, F. & Monserrat, V. J., 1991b. Contribución al conocimiento de los neurópteros de Albacete (Insecta, Planipennia). Jornadas Sobre el Medio Natural Albacetense, Albacete, 1990: 179-184.
|
| ○ |
Marín, F. & Monserrat, V. J., 1995a. Contribución al conocimiento de los Neurópteros de Valencia (Insecta, Neuroptera). Boletín de la Asociación española de Entomología, 19, 3-4: 35-49.
|
| ○ |
Marín, F. & Monserrat, V. J., 1995b. Contribución al conocimiento de los neurópteros de Zaragoza (Insecta, Neuropteroidea).
Zapateri, Revista Aragonesa de Entomología, 5: 109-126.
|
| ○ |
Martín Albaladejo, C., 1994. Bibliografía entomológica de autores españoles (1758-1990). Documentos Fauna Ibérica, 1. Museo Nacional de Ciencias Naturales. Madrid. 821 pp.
|
| ○ |
Martins, C. C. & Amorim, D. S., 2016. Brazilian dustywings (Neuroptera: Coniopterygidae): new species of Incasemidalis Meinander, 1972 and Coniopteryx Curtis, 1834, checklist and key for the Brazilian species. Zootaxa, 4083(2): 257-289. http://dx.doi.org/10.11646/zootaxa.4083.2.6 |
| ○ |
Martins, F. M., Mendoça, T. R., Lavadinho, A. M. P. & Vieira, M. M., 2002. Entomofauna num pomar de limoeiros, no Escaroupim
(Ribatejo), em Portugal. Boletín de Sanidad Vegetal, Plagas, 28: 435-443.
|
| ○ |
McEwen, P. K., New, T. R. & Whittington, A. E., 2001. Lacewings in the Crop Environment. Cambridge University Press. Cambridge. 546 pp.
|
| ○ |
McEwen, P. K. & Oswald, J. D., 1998. Neuroptera on the Internet. In: Panelius, S. P. (ed.). Neuropterology 1997 (Proceedings of the Sixth International Symposium on Neuropterology, Helsinki 1997). Acta Zoologica Fennica, 209: 151-152.
|
| ○ |
McKellar, R., Wolfe, A. P., Tappert, R. & Muehlenbachs, K., 2008. Correlation of Grassy Lake and Cedar Lake ambers using infrared
spectroscopy, stable isotopes, and palaeoentomology. Canadian Journal of Earth Sciences, 45(9): 1061-1082.
|
| ○ |
McLachlan, R., 1867. New genera and species, & c., of neuropterous insects; and a revision of Mr. F. Walker’s British Museum
Catalogue of Neuroptera, part ii. (1853), as far as the end of the genus Myrmeleon. Journal of the Linnean Society of London, Zoology, 9: 230-281.
|
| ○ |
McLachlan, R., 1868. A monograph of the British Neuroptera-Planipennia. Transactions of the Royal Entomological Society of London, 16: 145-224. http://dx.doi.org/10.1111/j.1365-2311.1868.tb00622.x |
| ○ |
McLachlan, R., 1870a. A catalogue of British Neuroptera. Entomological Society of London. London. VIII + 42 pp.
|
| ○ |
McLachlan, R., 1870b. The sexes of Coniopteryx psociformis. The Entomologist’s Monthly Magazine, 6: 238.
|
| ○ |
McLachlan, R., 1880. Notes on the entomology of Portugal. II. Pseudo-Neuroptera (in part) & Neuroptera-Planipennia. The Entomologist’s Monthly Magazine, 17: 103-108.
|
| ○ |
McLachlan, R., 1882a. The Neuroptera of Madeira and the Canary Islands. Journal of the Linnean Society of London, Zoology, 16: 149-183.
|
| ○ |
McLachlan, R., 1882b. Nota [Note on some Neuroptera from Switzerland and north Italy]. Transactions of the Royal Entomological Society of London (Proceedings), 30: xviii. http://dx.doi.org/10.1111/j.1365-2311.1882.tb02035.x |
| ○ |
McLachlan, R., 1882c. Nota [Note on some Neuroptera from Switzerland and north Italy]. The Entomologist’s Monthly Magazine, 19: 143.
|
| ○ |
Meinander, M., 1962a. The Neuroptera and Mecoptera of eastern Fennoscandia. Fauna Fennica, 13: 1-96.
|
| ○ |
Meinander, M., 1962b. Some Neuroptera from the Madeira and Canary Islands. Notulae Entomologicae, 42: 79-82.
|
| ○ |
Meinander, M., 1963. Coniopterygidae (Neuroptera) from Morocco. Notulae Entomologicae, 43: 92-164.
|
| ○ |
Meinander, M., 1965a. Coniopterygidae (Neuroptera) from the Sudan. Notulae Entomologicae, 45: 65-72.
|
| ○ |
Meinander, M., 1965b. Some Neuroptera from the Canary Islands and the Spanish Sahara. Notulae Entomologicae, 45: 53-60.
|
| ○ |
Meinander, M., 1969a. The genus Heteroconis Enderlein, 1905 (Neuroptera, Coniopterygidae). Notulae Entomologicae, 49: 49-68.
|
| ○ |
Meinander, M., 1969b. Coniopterygidae from Mongolia (Neuroptera). Notulae Entomologicae, 49: 7-10.
|
| ○ |
Meinander, M., 1971. Coniopterygidae from Mongolia II (Neuroptera). Notulae Entomologicae, 51: 41-44.
|
| ○ |
Meinander, M., 1972a. A Revision of the family Coniopterygidae (Planipennia). Acta Zoologica Fennica, 136: 1-357.
|
| ○ |
Meinander, M., 1972b. Coniopterygidae from Mongolia III (Neuroptera). Notulae Entomologicae, 52: 127-138.
|
| ○ |
Meinander, M., 1972c. The invertebrate fauna of the Kilpisjärvi area, Finnish Lapland. 8. Neuroptera and Mecoptera. Acta Societatis pro Fauna et Flora Fennica, 80: 93-98.
|
| ○ |
Meinander, M., 1973. Notes on some types of Enderlein’s species of Coniopterygidae, with description of a new species of Pampoconis (Neuroptera). Notulae Entomologicae, 53: 23-25.
|
| ○ |
Meinander, M., 1974a. The larvae of two North American species of Coniopterygidae (Neuroptera). Notulae Entomologicae, 54:12-16.
|
| ○ |
Meinander, M., 1974b. Coniopterygidae from Madagascar (Neuroptera). Notulae Entomologicae, 54: 60-63.
|
| ○ |
Meinander, M., 1974c. Coniopterygidae from South and Central America (Neuroptera). Notulae Entomologicae, 54: 97-106.
|
| ○ |
Meinander, M., 1974d. Coniopterygidae from western North America (Neuroptera). Entomologica Scandinavica, 5(3): 217-232. http://dx.doi.org/10.1163/187631274X00254 |
| ○ |
Meinander, M., 1975a. Fossil Coniopterygidae (Neuroptera). Notulae Entomologicae, 55: 53-57.
|
| ○ |
Meinander, M., 1975b. Coniopterygidae from West Africa (Insecta: Neuroptera). Entomologica Scandinavica, 6: 247-252. http://dx.doi.org/10.1163/187631275X00082 |
| ○ |
Meinander, M., 1975c. Coniopterygidae from North America (Neuroptera). Notulae Entomologicae, 55: 28-32.
|
| ○ |
Meinander, M., 1975d. Coniopterygidae from South Africa (Neuroptera). Notulae Entomologicae, 55: 81-85.
|
| ○ |
Meinander, M., 1976. Coniopterygidae from Africa (Neuroptera). Notulae Entomologicae, 56: 85-88.
|
| ○ |
Meinander, M., 1977. Coniopterygidae from the Arabian Peninsula (Neuroptera). Entomologica Scandinavica, 8: 81-85. http://dx.doi.org/10.1163/187631277X00143 |
| ○ |
Meinander, M., 1979a. The phylogeny and geographical distribution of the Aleuropteryginae (Neuroptera, Coniopterygidae). Annales Entomologici Fennici, 45: 16-23.
|
| ○ |
Meinander, M., 1979b. Insects of Saudi Arabia. Neuroptera: Fam. Coniopterygidae. Fauna of Saudi Arabia, 1: 334-341.
|
| ○ |
Meinander, M., 1980. Coniopterygidae from Brazil (Neuroptera). Entomologica Scandinavica, 11: 129-144. http://dx.doi.org/10.1163/187631280X00455 |
| ○ |
Meinander, M., 1981. A review of the genus Coniopteryx (Neuroptera,Coniopterygidae). Annales Entomologici Fennici, 47: 97-110.
|
| ○ |
Meinander, M., 1982. The Coniopterygidae of Ceylon (Neuroptera). Entomologica Scandinavica, 13: 49-55. http://dx.doi.org/10.1163/187631282X00561 |
| ○ |
Meinander, M., 1983a. Coniopterygidae from South and Central America II (Neuroptera). Neuroptera International, 2: 179-198.
|
| ○ |
Meinander, M., 1983b. The Coniopterygidae (Neuroptera) of southern Africa and adjacent Indian Ocean Islands. Annals of the Natal Museum, 25(2): 475-499.
|
| ○ |
Meinander, M., 1986. The Coniopterygidae of America (Neuroptera). In: Gepp, J., Aspöck, H. & Hölzel, H. (eds.). Recent Research in Neuropterology (Proceedings of the 2nd International Symposium on Neuropterology, Hamburg 1984). Graz: 31-43.
|
| ○ |
Meinander, M., 1990. The Coniopterygidae (Neuroptera, Planipennia). A check list of the species of the world, descriptions
of new species and other new data. Acta Zoologica Fennica, 189: 1-95.
|
| ○ |
Meinander, M., 1992. A review of the family Coniopterygidae (Insecta: Neuroptera). In: Canard, M., Aspöck, H. & Mansell, M.
W. (eds.). Current Research in Neuropterology (Proceedings of the Fourth International Symposium on Neuropterology, Bagnères-de-Luchon
1991). Toulouse: 255-260.
|
| ○ |
Meinander, M., 1995. Coniopterygidae (Neuroptera, Planipennia) from Costa Rica and Nicaragua. Brenesia, 43/44: 61-70.
|
| ○ |
Meinander, M., 1996. Coniopterygidae from sub-Saharan Africa (Insect: Neuroptera). In: Canard, M., Aspöck, H. & Mansell, M.
W. (eds.). Current Research in Neuropterology (Proceedings of the Fourth International Symposium on Neuropterology, Bagnères-de-Luchon
1991). Toulouse: 187-192.
|
| ○ |
Meinander, M., 1998a. Coniopterygidae (Neuroptera) from the Mediterranean Region and Iran. Journal of Neuropterology, 1: 23-31.
|
| ○ |
Meinander, M., 1998b. Coniopterygidae (Neuroptera) from southern and eastern Africa. African Entomology, 6(1): 117-146.
|
| ○ |
Meinander, M., 1998c. Coniopterygidae (Neuroptera) in amber from the Dominican Republic. Journal of Neuropterology, 1: 33-36.
|
| ○ |
Meinander, M., 2002. Family Coniopterygidae. In: Penny, N. D. (ed.). A Guide to the Lacewings (Neuroptera) of Costa Rica. Proceedings of the California Academy of Sciences, (4), 53: 227-236 (texto), 374-393 (figuras).
|
| ○ |
Meinander, M. & Penny, N. D., 1982. Neuroptera of the Amazon Basin. Part 5. Coniopterygidae. Acta Amazonica, 12: 185-208.
|
| ○ |
Messner, B., 1965. Helicoconis aptera n. spec., ein neuer Vertreter der Unterfamilie Aleuropteryginae (Planipennia, Coniopterygidae) aus Bulgarien. Deutsche Entomologische Zeitschrift (N.F.), 12: 337-340.
|
| ○ |
Meunier, F. [A.], 1910a. Un Coniopterygidae du copal de récent de Madagascar [Nevr.]. Bulletin de la Société Entomologique de France, 1910: 164-166.
|
| ○ |
Meunier, F. [A.], 1910b. Un Coniopterygidae du copal récent de Togo (Afrique) [Nevr.]. Bulletin de la Société Entomologique de France, 1910: 119-120.
|
| ○ |
Meunier, F. [A.], 1910c. Un Coniopterygidae du copal récent de Togo. Annales de la Société Scientifique de Bruxelles, 34: 198-199.
|
| ○ |
Miller, G. L., Oswald, J. D. & Miller, D. R., 2004. Lacewings and scale insects: a review of predator/prey associations between
the Neuropterida and Coccoidea (Insecta: Neuroptera, Raphidioptera, Hemiptera). Annals of the Entomological Society of America, 97: 1103-1125. http://dx.doi.org/10.1603/0013-8746(2004)097%5B1103:LASIAR%5D2.0.CO;2
|
| ○ |
Mjöberg, E. [G.], 1909. Svensk insektfauna. 8. Neuroptera. I. Första Underordningen Planipennia. Entomologisk Tidskrift, 30: 129-161.
|
| ○ |
Monserrat, V. J., 1975. Descripción de una nueva especie del género Coniopteryx Curtis 1834 en España (Neuroptera: Planipennia). Vie et Milieu. Série C, Biologie Terrestre, 25: 259-265.
|
| ○ |
Monserrat, V. J., 1976a. Un nuevo coniopterígido en España. Nouvelle Revue d’Entomologie, 6(1): 97-100.
|
| ○ |
Monserrat, V. J., 1976b. Hemisemidalis pallida Withycombe, Coniopteryx atlasensis Meinander dos nuevos Neuropteros para la fauna europea (Planipennia). Vie et Milieu. Série C, Biologie Terrestre, 26: 247-251.
|
| ○ |
Monserrat, V. J., 1977a. Los Neurópteros del Guadarrama. Trabajos de la Cátedra de Entomología, Facultad de Biología, Universidad Complutense de Madrid, 17: 1-57.
|
| ○ |
Monserrat, V. J., 1977b. Descripción de una nueva especie del género Aleuropteryx Löw 1885 en la Península Ibérica (Neuroptera Planipennia). Annali del Museo Civico di Storia Naturale Giacomo Doria, 81: 365-373.
|
| ○ |
Monserrat, V. J., 1977c. Tres nuevas especies del género Coniopteryx Curtis, 1834, para la fauna española. Miscelánea Zoológica, 4: 139-142.
|
| ○ |
Monserrat, V. J., 1978a. Primera contribución al conocimiento de los Neurópteros de Cádiz (Insecta, Neuroptera, Planipennia).
Boletín de la Real Sociedad Española de Historia Natural, Sección Biológica, 76: 57-70.
|
| ○ |
Monserrat, V. J., 1978b. Contribución al conocimiento de los Neurópteros de Orense (Neu. Planipennia). Boletín de la Asociación Española de Entomología, 2: 169-184.
|
| ○ |
Monserrat, V. J., 1978c. Sobre los Neuropteros de las Islas Canarias II: Semidalis candida Navás, 1916 (Neur. Coniopterygidae). Nouvelle Revue d’Entomologie, 8: 369-376.
|
| ○ |
Monserrat, V. J., 1978d. Redescripción de Semidalis palmensis (Klingstedt, 1936) (Neu. Plan., Coniopterygidae). Boletín de la Asociación Española de Entomología, 1: 141-150.
|
| ○ |
Monserrat, V. J., 1979a. Segunda contribución al conocimiento de los neurópteros de Cádiz (Insecta, Planipennia). Boletín de la Real Sociedad Española de Historia Natural, Sección Biológica, 77: 409-417.
|
| ○ |
Monserrat, V. J., 1979b. Aportación al conocimiento de los Neurópteros (Insecta, Planipennia) de las Islas de El Hierro y
de La Gomera (Canarias). Boletín de la Asociación Española de Entomología, 3: 79-83.
|
| ○ |
Monserrat, V. J., 1979c. Coniopteryx arcuata Kis, 1965, nueva para la fauna Española (Neuropt. Coniopterygidae). Graellsia, 33: 103-106.
|
| ○ |
Monserrat, V. J., 1980. Contribución al conocimiento de los Neurópteros de Toledo. Graellsia, [1978], 34: 177-193.
|
| ○ |
Monserrat, V. J., 1981a. Dos nuevos coniopterígidos de Paraguay (Insecta, Neuroptera, Coniopterygidae). Eos, 55/56: 155-163.
|
| ○ |
Monserrat, V. J., 1981b. Contribucion al conocimiento de los Neurópteros de Italia (Neuroptera, Planipennia). Neuroptera International, 1: 48-64.
|
| ○ |
Monserrat, V. J., 1981c. Coniopteryx (Holoconiopteryx) haematica McLachlan, 1868, (Neur. Plan., Coniopterygidae), nueva para la fauna Ibérica. Boletín de la Asociación Española de Entomología, 4: 179-183.
|
| ○ |
Monserrat, V. J., 1982a. Contribución al conocimiento de los Neurópteros de Cáceres. Graellsia, 38: 67-84.
|
| ○ |
Monserrat, V. J., 1982b. Coniopteryx (Xeroconiopteryx) kerzhneri Meinander, 1971 (Neur., Plan., Coniopterygidae) nueva especie para la fauna europea. Boletín de la Asociación Española de Entomología, 6: 57-61.
|
| ○ |
Monserrat, V. J., 1982c. Contribución al conocimiento de los coniopterígidos (Insecta, Neuroptera, Coniopterygidae) de la
región oriental. Annali del Museo Civico di Storia Naturale Giacomo Doria, 84: 9-39.
|
| ○ |
Monserrat, V. J., 1983a. Contribución al conocimiento de los Coniopterígidos de Venezuela. (Neuroptera, Planipennia, Coniopterygidae).
Boletín de Entomología Venezolana (N.S.), 2: 137-154.
|
| ○ |
Monserrat, V. J., 1983b. Pterocroce capillaris (Klug, 1836) en Europa (Neur., Plan., Nemopteridae). Neuroptera International, 2, 3: 109-128.
|
| ○ |
Monserrat, V. J., 1984a. Correcciones a las citas de Coniopterígidos (Insecta, Planipennia, Coniopterygidae) dadas por L.
Navás. Miscel·lània Zoològica, 8: 145-151.
|
| ○ |
Monserrat, V. J., 1984b. Contribución al conocimiento de los neurópteros de Alicante (Neur., Planipennia). Mediterránea (Serie de Estudios Biológicos), 7: 91-116.
|
| ○ |
Monserrat, V. J., 1984c. Contribución al conocimiento de los neurópteros de Zamora (Neur., Planipennia). Miscel·lània Zoològica, 8: 153-163.
|
| ○ |
Monserrat, V. J., 1984d. Contribución al conocimiento de los neurópteros de Huesca (Neur., Planipennia). Pirineos, 121: 29-50.
|
| ○ |
Monserrat, V. J., 1984e. Contribución al conocimiento de los neurópteros de Salamanca (Neur., Planipennia). Eos, [1983], 59: 165-177.
|
| ○ |
Monserrat, V. J., 1984f. Contribución al conocimiento de los Coniopterígidos de México (Neuroptera, Planipennia, Coniopterygidae).
Eos, 60: 211-222.
|
| ○ |
Monserrat, V. J., 1984g. Nuevos datos sobre los Coniopterígidos (Neur., Plan. Coniopterygidae) ibéricos. Boletín de la Asociación Española de Entomología, 8: 25-34.
|
| ○ |
Monserrat, V. J., 1985a. Contribución al conocimiento de los neurópteros de Lugo (Neur., Planipennia). Trabajos Compostelanos de Biología, 12: 87-98.
|
| ○ |
Monserrat, V. J., 1985b. Contribución al conocimiento de los Neurópteros (Neuroptera: Planipennia) de Murcia. Anales de Biología (Biología Animal), 1: 81-94.
|
| ○ |
Monserrat, V. J., 1985c. Lista de los tipos de Mecoptera y Neuroptera (Insecta) de la colección L. Navás, depositados en el
Museo de Zoología de Barcelona. Miscel·lània Zoològica, 9: 233-243.
|
| ○ |
Monserrat, V. J., 1985d. Nuevos datos sobre los Coniopterígidos (Neur., Plan., Coniopterygidae) ibéricos. Boletín de la Asociación Española de Entomología, 9: 127-141.
|
| ○ |
Monserrat, V. J., 1985e. Contribución al conocimiento de los neurópteros de Marruecos (Insecta, Neuroptera, Planipennia).
Mediterránea (Serie de Estudios Biológicos), 8: 73-82.
|
| ○ |
Monserrat, V. J., 1986. Longinos Navás, his neuropterological work and collection. In: Gepp, J., Aspöck, H. & Hölzel, H. (eds.).
Recent Research in Neuropterology (Proceedings of the 2nd International Symposium on Neuropterology, Hamburg 1984). Graz: 173-176.
|
| ○ |
Monserrat, V. J., 1987a. Contribución al conocimiento de los Neurópteros de Almería (Neur., Planipennia). Graellsia, [1986], 42: 131-147.
|
| ○ |
Monserrat, V. J., 1987b. Els megalòpters, Els Rafidiòpters, Els Planipennis: crisopes, reis de formigues i afins. Història Natural dels Països Catalans, 10(2): 251-265.
|
| ○ |
Monserrat, V. J., 1989a. Contribución al conocimiento de los Coniopterígidos de Rio Muni (Guinea Ecuatorial) (Neuropteroidea,
Planipennia: Coniopterygidae). Annali del Museo Civico di Storia Naturale di Genova, 87: 157-181.
|
| ○ |
Monserrat, V. J., 1989b. Algunos Neurópteros del Museo de Basilea (Insecta, Neuropteroidea, Planipennia). Entomologica Basiliensia, 13: 417-428.
|
| ○ |
Monserrat, V. J., 1995. Nuevos datos sobre los coniopterígidos de las regiones paleártica y afrotropical. (Neuroptera: Coniopterygidae).
Graellsia, [1994], 50: 109-127.
|
| ○ |
Monserrat, V. J., 1996a. Lista de los neurópteros de Aragón (Megaloptera, Raphidioptera, Planipennia). Catalogus de la entomofauna Aragonesa, 11: 11-17.
|
| ○ |
Monserrat, V. J., 1996b. Nuevos datos sobre los coniopterígidos de Yemen (Neuroptera: Coniopterygidae). Annali del Museo Civico di Storia Naturale di Genova, 591: 1-26.
|
| ○ |
Monserrat, V. J., 1998. Nuevos datos sobre los coniopterígidos de Zimbabwe y Zambia (Neuroptera: Coniopterygidae). Journal of Neuropterology, 1: 5-15.
|
| ○ |
Monserrat, V. J., 2001. Recognition of larval Neuroptera. Coniopterygidae. In: McEwen, P. K., New, T. R. & Whittington, A.
E. (eds.). Lacewings in the Crop Environment. Cambridge University Press. Cambridge: 43-50.
|
| ○ |
Monserrat, V. J., 2002. New data on the dusty wings from Africa and Europe (Insecta, Neuroptera: Coniopterygidae). Graellsia, 58(1): 3-19. http://dx.doi.org/10.3989/graellsia.2002.v58.i1.263 |
| ○ |
Monserrat, V. J., 2005a. Catálogo de los Neurópteros de Baleares con nuevos datos sobre su fauna (Insecta, Neuroptera). Bolletí de la Societat d′Història Natural de les Balears, 48: 71-85.
|
| ○ |
Monserrat, V. J., 2005b. New data on the dusty wings from Coquimbo, Patagonia and Tierra del Fuego (Neuroptera, Coniopterygidae).
Entomofauna, 25: 421-433.
|
| ○ |
Monserrat, V. J., 2006. New taxonomical and faunistic data sobre the dusty wings from Senegal (Insecta, Neuroptera: Coniopterygidae).
Graellsia, 62(1): 13-24. http://dx.doi.org/10.3989/graellsia.2006.v62.i1.25 |
| ○ |
Monserrat, V. J., 2010. Nuevas o interesantes citas de neurópteros en la Península Ibérica (Insecta: Neuroptera). Heteropterus Revista de Entomología, 10(1): 19-34.
|
| ○ |
Monserrat, V. J., 2011. Sobre algunas especies de neurópteros de la Península Ibérica y de las Islas Canarias de posición
taxonómica problemática o con citas dudosas o cuestionables (Insecta, Neuroptera: Megaloptera, Planipennia). Boletín de la Sociedad Entomológica Aragonesa, 49: 153-178.
|
| ○ |
Monserrat, V. J., 2013. Los Neurópteros (Neuroptera). In: Ruano, F., Tierno de Figueroa, M. y Tinaut, A. (eds). Los Insectos de Sierra Nevada. 200 años de historia. Asociación española de Entomología. Granada: 281-309.
|
| ○ |
Monserrat, V. J., 2015. Los hemeróbidos de la Península Ibérica y Baleares (Insecta, Neuropterida, Neuroptera: Hemerobiidae).
Graellsia, 71(2): e02. http://dx.doi.org/10.3989/graellsia.2015.v71.129 |
| ○ |
Monserrat, V. J. & Bayo, J., 1995. Nuevos datos sobre los coniopterígidos de Méjico central (Neuroptera: Coniopterygidae).
Boletín de la Asociación Española de Entomología, 19: 125-130.
|
| ○ |
Monserrat, V. J. & Díaz-Aranda, L. M., 1987. Contribución al conocimiento de los neurópteros (Neur.: Raphidioptera, Planipennia)
de Cuenca. Boletín de la Asociación Española de Entomología, 11: 171-189.
|
| ○ |
Monserrat, V. J. & Díaz-Aranda, L. M., 1988a. Contribución al conocimiento de los Neurópteros de Castellón (Insecta, Neuropteroidea,
Planipennia). Mediterránea (Serie de Estudios Biológicos), 10: 87-95.
|
| ○ |
Monserrat, V. J. & Díaz-Aranda L. M., 1988b. Contribución al conocimiento de los coniopterígidos de la Isla de Bioco (Guinea
Ecuatorial) (Neuropteroidea, Planipennia: Coniopterygidae). Revue de Zoologie Africaine, 102: 493-502.
|
| ○ |
Monserrat, V. J. & Díaz-Aranda, L. M., 1989. Contribución al conocimiento de los Neurópteros de Castellón (Insecta, Neuropteroidea,
Planipennia). Mediterránea (Serie de Estudios Biológicos), 11: 47-58.
|
| ○ |
Monserrat, V. J. & Díaz-Aranda L.M., 1990. Nuevos datos sobre los coniopterígidos ibéricos (Neuroptera: Coniopterygidae).
Neuroptera International, 6(1): 39-49.
|
| ○ |
Monserrat, V. J., Díaz-Aranda, L. M. & Hölzel, H., 1991. Contribución al conocimiento de los neurópteros de Marruecos (Insecta,
Neuropteroidea). Eos, 66: 101-115.
|
| ○ |
Monserrat, V. J. & El-Serwy, S.A., 2010. Nimboa adelae Monserrat (Neuroptera: Coniopterygidae), a newly recorded predator on the red-striped soft scale, Pulvinaria tenuivalvata (Newstead), (Hemiptera: Coccidae) in Egypt. XII International Symposium on Scale Insect Studies, Chania: 57.
|
| ○ |
Monserrat, V. J., Gamarra, P. & Deretsky, Z., 1999. The neuropterological fauna of the houses of the city of Madrid. Journal of Neuropterology, 2: 35-44.
|
| ○ |
Monserrat, V. J. & Hölzel, H., 1987. Contribución al conocimiento de los neurópteros de Anatolia (Neuropteroidea, Planipennia).
Eos, 63: 133-142.
|
| ○ |
Monserrat, V. J. & Marín, F., 1992. Substrate specificity of Iberian Coniopterygidae (Insecta: Neuroptera). In: Canard, M.,
Aspöck, H. & Mansell, M. W. (eds.). Current Research in Neuropterology (Proceedings of the Fourth International Symposium on Neuropterology, Bagnères-de-Luchon
1991). Toulouse: 279-290.
|
| ○ |
Monserrat, V. J. & Marín, F., 2001. Comparative plant substrate specificity of Iberian Hemerobiidae, Coniopterygidae and Chrysopidae.
In: McEwen, P. K., New, T. R. & Whittington, A. E. (eds.). Lacewings in the Crop Environment. Cambridge University Press. Cambridge: 424-434.
|
| ○ |
Monserrat, V. J., Marín, F. & Díaz-Aranda, L. M., 1994. Contribución al conocimiento de los neurópteros de Lérida (Insecta,
Neuroptera). Zoologixa Baetica, 5: 41-64.
|
| ○ |
Monserrat, V. J., Oswald, J.D., Tauber, C. A. & Díaz-Aranda, L. M., 2001. Recognition of larval Neuroptera. In: McEwen, P.
K., New, T. R. & Whittington, A. E. (eds.). Lacewings in the Crop Environment. Cambridge University Press. Cambridge: 43-81.
|
| ○ |
Monserrat, V. J. & Reviejo, I., 1977. Aportación al conocimiento de los Neurópteros (Insecta, Planipennia) de la Isla Tenerife.
Nouvelle Revue d’Entomologie, 7: 351-354.
|
| ○ |
Monserrat, V. J. & Reviejo, I., 1978. Aportación al conocimiento de los Neurópteros (Insecta, Planipennia) de la Isla de la
Palma. Nouvelle Revue d’Entomologie, 8: 363-368.
|
| ○ |
Monserrat, V. J. & Reviejo, I., 1980. Sobre los Neurópteros de Túnez, I: Coniopterygidae (Neuroptera, Planipennia). Nouvelle Revue d’Entomologie, 10: 359-361.
|
| ○ |
Monserrat, V. J. & Triviño, V., 2013. Atlas de los neurópteros de la Península Ibérica e Islas Baleares (Insecta, Neuroptera: Megaloptera, Raphidioptera, Planipennia)
/ Atlas of the Iberian and Balearic lacewings (Insecta, Neuroptera: Megaloptera, Raphidioptera, Planipennia). Monografías de la Sociedad Entomológica Aragonesa, vol. 13. Zaragoza. 154 pp., 136 mapas.
|
| ○ |
Monserrat, V. J., Triviño, V. & Acevedo, F., 2013. Contribución al conocimiento de los neurópteros de Navarra (Insecta, Neuroptera).
Heteropterus Revista de Entomología, 13(1): 41-58.
|
| ○ |
Morgan, M. J., 1980. Contribución al catálogo faunístico del Coto Nacional de las Sierras de Cazorla y Segura. Neuroptera.
In: M.G. de Viedma (Ed.). Fauna de Cazorla. Invertebrados. Monografías Ministerio Agricultura, Monografías ICONA, 23. Madrid: 119.
|
| ○ |
Morinière, J., Hendrich, L., Hausmann, A., Hebert, P., Haszprunar, G. & Gruppe, A., 2014. Barcoding Fauna Bavarica: 78% of
the Neuropterida Fauna Barcoded! PLoS ONE 9(10): e109719. http://dx.doi.org/10.1371/journal.pone.0109719 |
| ○ |
Muma, M. H., 1967. Biological notes on Coniopteryx vicina (Neuroptera: Coniopterygidae). The Florida Entomologist, 50: 285-293. http://dx.doi.org/10.2307/3493158 |
| ○ |
Muma, M. H., 1971. Coniopterygids (Neuroptera) on Florida citrus trees. The Florida Entomologist, 54: 283-288. http://dx.doi.org/10.2307/3493587 |
| ○ |
Murphy, D. H. & Lee, Y. T., 1971. Three new species of Coniopteryx from Singapore (Plannipennia [sic]: Coniopterygidae). Journal of Entomology Series B, Taxonomy, 40: 151-161. http://dx.doi.org/10.1111/j.1365-3113.1971.tb00118.x |
| ○ |
Najarro, M., Peñalver, E., Pérez-de la Fuente, R., Ortega-Blanco, J., Menor-Salván, C., Barrón, E., Soriano, C., Rosales,
I., López del Valle, R., Velasco, F., Tornos, F., Daviero-Gomez, V., Gomez, B. & Delclòs, X., 2010. Review of the El Soplao
amber outcrop, Early Cretaceous of Cantabria, Spain. Acta Geologica Sinica (English Edition), 84: 959-976.
|
| ○ |
Nakahara, W., 1913. [Studies on Japanese Coniopterygidae.] Zoological Magazine, 25: 195-201. (en japonés).
|
| ○ |
Narayanan, E. S., 1942. On the bionomics and life-history of Coniopteryx pusana Withycombe Coniopterygidae (Neuroptera). Indian Journal of Entomology, 4: 1-4.
|
| ○ |
Navás, L., 1905a. Notas entomológicas. XII. Neurópteros de Pozuelo de Calatrava (C. Real). Boletín de la Real Sociedad Española de Historia Natural, 5: 503-509.
|
| ○ |
Navás, L., 1905b. Catálogo descriptivo de los insectos Neurópteros de los alrededores de Madrid. Revista de la Real Academia de Ciencias Exactas Fisicas y Naturales de Madrid, 2: 521-574.
|
| ○ |
Navás, L., 1906. Catálogo descriptivo de los insectos Neurópteros de las Islas Canarias. Revista de la Real Academia de Ciencias Exactas Fisicas y Naturales de Madrid, 4: 687-706.
|
| ○ |
Navás, L., 1908. Neurópteros de España y Portugal [part 3]. Brotéria (Zoológica), 7: 5-131.
|
| ○ |
Navás, L., 1910a. Mis excursiones entomológicas durante el verano de 1909 (2 Julio-3 Agosto). Butlletí de la Institució Catalana d’Història Natural, 10: 32-56, 74-75.
|
| ○ |
Navás, L., 1910b. Notas entomológicas (2. Serie). 2. Excursiones por Cataluña y Mallorca. Boletín de la Sociedad Aragonesa de Ciencias Naturales, 9: 240-248.
|
| ○ |
Navás, L., 1911. Excursiones por los alrededores de Granada. Boletín de la Sociedad Aragonesa de Ciencias Naturales, 10: 204-211.
|
| ○ |
Navás, L., 1912. Insectos Neurópteros nuevos. Verhandlungen des VIII Internationalen Zoologen-kongresses, Graz 1910. Gustav Fischer. Jena: 746-751.
|
| ○ |
Navás, L., 1913a. Notas entomológicas, 2.a serie. 5. Cuatro pequeñas colecciones de Neurópteros de la península Ibérica. Boletín de la Sociedad Aragonesa de Ciencias Naturales, 12: 77-89.
|
| ○ |
Navás, L., 1913b. Excursiones por los alrededores de Zaragoza. Boletín de la Sociedad Aragonesa de Ciencias Naturales, 12: 61-69, 75-77.
|
| ○ |
Navás, L., 1914a. Excursión anual de la Sociedad Aragonesa de Ciencias Naturales a Ortigosa y Valvanera (Logroño). Conclusión.
Boletín de la Sociedad Aragonesa de Ciencias Naturales, 13: 29-37.
|
| ○ |
Navás, L., 1914b. Neurópteros del Moncayo (Zaragoza). Boletín de la Sociedad Aragonesa de Ciencias Naturales, 13: 207-218.
|
| ○ |
Navás, L., 1914c. Algunos neurópteros de los alrededores de Madrid. Boletín de la Sociedad Aragonesa de Ciencias Naturales, 13: 38-41.
|
| ○ |
Navás, L., 1914d. Notas entomológicas. 10. Neurópteros de Mallorca. Boletín de la Sociedad Aragonesa de Ciencias Naturales, 13, 9: 185-192.
|
| ○ |
Navás, L., 1915. Notas entomológicas, 2a Serie. Excursiones por Cataluña. Julio 1914. Boletín de la Sociedad Aragonesa de Ciencias Naturales, 14: 27-32, 35-59, 66-80.
|
| ○ |
Navás, L., 1916a. Excursiones por el norte de España. Congreso de la Asociación Española para el Progreso de las Ciencias, 6: 161-179.
|
| ○ |
Navás, L., 1916b. Excursions entomològiques al nort de la provincia de Lleida. Butlletí de la Institució Catalana d’Història Natural, 16: 150-158.
|
| ○ |
Navás, L., 1916c. Excursión al valle de Arán (Lerida) 17-28 de Julio de 1915. Boletín de la Sociedad Aragonesa de Ciencias Naturales, 15: 179-194.
|
| ○ |
Navás, L., 1916d. Neuroptera nova Africana. VII Series. Memorie dell’Accademia Pontifica dei Nuovi Lincei, (2), 2: 51-58.
|
| ○ |
Navás, L., 1917a. Notas entomológicas. 2a serie. Neurópteros de Andorra. Boletín de la Sociedad Aragonesa de Ciencias Naturales, 17: 36-46.
|
| ○ |
Navás, L., 1917b. Comunicaciones entomológicas. 2. Excursiones entomológicas por Aragón y Navarra. Revista de la Real Academia de Ciencias Exactas Físico-Químicas y Naturales de Zaragoza, 2: 81-91.
|
| ○ |
Navás, L., 1918a. Notas entomológicas. Neurópteros de Andorra (Suplemento). Boletín de la Sociedad Aragonesa de Ciencias Naturales, 17: 231-235.
|
| ○ |
Navás, L., 1918b. Insecta nova. IV Series. Memorie dell’Accademia Pontifica dei Nuovi Lincei, 4: 13-23.
|
| ○ |
Navás, L., 1918c. Excursiones entomológicas por el norte de la provincia de Lérida. Butlletí de la Institució Catalana d’Història Natural, 18: 36-49.
|
| ○ |
Navás, L., 1919a. Excursiones entomológicas por Cataluña durante el verano de 1918. Memorias de la Real Academia de Ciencias y Artes de Barcelona, (3), 15: 181-214.
|
| ○ |
Navás, L., 1919b. Notas entomológicas, 2a serie. Excursión anual de la Sociedad Aragonesa de Ciencias Naturales al Valle de Ordesa (Huesca) (22-29 de julio de 1918).
Boletín de la Sociedad Ibérica de Ciencias Naturales, 18: 37-44.
|
| ○ |
Navás, L., 1921a. Mis excursiones científicas del verano de 1919. Memorias de la Real Academia de Ciencias y Artes de Barcelona, (3), 17: 143-169.
|
| ○ |
Navás, L., 1921b. Excursiones científicas realizadas durante el verano de 1920. Asociación Española para el Progreso de las Ciencias. Congreso de Oporto, 4: 59-74.
|
| ○ |
Navás, L., 1922. Mis excursiones entomológicas durante el verano de 1921. Boletín de la Sociedad Entomologica de España, 5: 107-119.
|
| ○ |
Navás, L., 1923a. Excursiones por Aragón durante el verano de 1923. Boletín de la Sociedad Ibérica de Ciencias Naturales, 22: 161-176.
|
| ○ |
Navás, L., 1923b. Excursions entomològiques de l’istiu de 1922 (26 de juny-26 de juliol). Arxius de l’Institut de Ciències, 8: 1-34.
|
| ○ |
Navás, L., 1924a. Entomologia de Catalunya. Neuròpters. Fasc. I. Neuròpters propis. Fauna de Catalunya, Institut d´Estudis Catalans. Barcelona. 271 pp.
|
| ○ |
Navás, L., 1924b. Excursió entomológica al Cabrerès (Girona-Barcelona). Musei Barcinonensis Scientiarum Naturalium Opera, 4(10): 1-59.
|
| ○ |
Navás, L., 1925a. Sinopsis de los Neurópteros (Ins.) de la Península Ibérica, Memorias de la Sociedad Ibérica de Ciencias Naturales, 4: 1-150.
|
| ○ |
Navás, L., 1925b. Mis excursiones del verano de 1925. Brotéria, 22: 131-140.
|
| ○ |
Navás, L., 1927a. Veinticinco formas nuevas de insectos. Boletín de la Sociedad Ibérica de Ciencias Naturales, 26 (9): 48-75.
|
| ○ |
Navás, L., 1927b. Mis excursiones científicas en 1926. Revista de la Real Academia de Ciencias Exactas Físico-Químicas y Naturales de Zaragoza, (1), 10: 81-124.
|
| ○ |
Navás, L., 1928. Comunicaciones entomológicas. 10. Mis excursiones científicas en 1927. Revista de la Real Academia de Ciencias Exactas Físico-Químicas y Naturales de Zaragoza, 11: 79-134.
|
| ○ |
Navás, L., 1929a. Excursiones por la provincia de Gerona en julio y agosto de 1928. Butlletí de la Institució Catalana d’Història Natural, 2(9): 27-48.
|
| ○ |
Navás, L., 1929b. Excursiones por Aragón durante el verano de 1929. Boletín de la Sociedad Entomologica de España, 12: 122-132.
|
| ○ |
Navás, L., 1930. Excursión a Ribera de Cardós, Pallars (Lérida). Butlletí de la Institució Catalana d’Història Natural, 2(10): 48-57.
|
| ○ |
Navás, L., 1931. De mis últimas excursiones entomológicas (1930-1931). Boletín de la Sociedad Entomologica de España, 14: 116-130.
|
| ○ |
Navás, L., 1932. Fáunula de Sobradiel (Zaragoza). Revista de la Real Academia de Ciencias Exactas Físico-Químicas y Naturales de Zaragoza, 16: 11-28.
|
| ○ |
Navás, L. & Marcet, A. F., 1910. Coniopterígido (Ins. Neur.) nuevo de Montserrat. Revista Montserratina, 4: 150-151.
|
| ○ |
Navone, P., 1987. Origine, struttura e funzioni di escreti e secreti entomatici di aspetto ceroso distribuiti sul corpo mediante
zampe. Annali della Facoltà di Scienze Agrarie della Università degli Studi di Torino, 14: 237-294.
|
| ○ |
Nel, A., 1991. Nouveaux insectes neuroptéroïdes fossiles de l’Oligocène de France (Neuroptera et Megaloptera). Bulletin du Muséum National d’Histoire Naturelle. Section C, Sciences de la Terre, Paléontologie, Géologie, Minérologie, 12: 327-349.
|
| ○ |
Nel, A., Perrichot, V. & Azar, D., 2005. New and poorly known fossil Coniopterygidae in Cretaceous and Cenozoic ambers (Insecta
: Neuroptera). Annales Zoologici, 55: 1-7.
|
| ○ |
Nelson, D. R., Freeman, T. P., Buckner, J. S., Hoelmer, K. A., Jackson, C. G. & Hagler, J. R., 2003. Characterization of the
cuticular surface wax pores and the waxy particles of the dustywing, Semidalis flinti (Neuroptera: Coniopterygidae). Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 136: 343-356. http://dx.doi.org/10.1016/S1096-4959(03)00216-1 |
| ○ |
New, T. R., 1967. Trap-banding as a collecting method for Neuroptera and their parasites, and some results obtained. Entomologist’s Gazette, 18: 37-44.
|
| ○ |
New, T. R., 1968. Dispersal of larvae of some British Neuroptera. Entomologist’s Gazette, 19: 230-232.
|
| ○ |
New, T. R., 1975. Lacewings (Neuroptera) as biological control agents. Victorian Entomologist, 5: 102-103.
|
| ○ |
New, T. R., 1986. A review of the Biology of Neuroptera Planipennia, Neuroptera International, Supplemental Series, 1: 1-58.
|
| ○ |
New, T. R., 1987. A new species of Heteroconis Enderlein (Neuroptera, Coniopterygidae) from Western Australia. Australian Entomological Magazine, 14: 1-3.
|
| ○ |
New, T. R., 1988. Coniopterygidae (Neuroptera) from the Wau area of Papua New Guinea. Neuroptera International, 5: 3-12.
|
| ○ |
New, T. R., 1989. Planipennia, Lacewings. Handbuch der Zoologie, 4(30): 1-132.
|
| ○ |
New, T. R., 1990. Coniopterygidae (Neuroptera) from Mt. Kinabalu, Sabah, East Malaysia. Neuroptera International, 6: 1-8.
|
| ○ |
New, T. R., 1991a. Insects as predators. University of New South Wales Press. 178 pp.
|
| ○ |
New, T. R., 1991b. Commentary: Prospects and priorities for conservation of Neuroptera. Neuroptera International, 6(4): 195-204.
|
| ○ |
New, T. R., 1998. Are Neuroptera an informative focal group for conservation assessment? In: Panelius, S. P. (ed.). Neuropterology 1997 (Proceedings of the Sixth International Symposium on Neuropterology, Helsinki 1997). Acta Zoologica Fennica, 209: 167-174.
|
| ○ |
New, T. R., 1999. Neuroptera and biological control (Neuropterida). Stapfia, 60: 147-166.
|
| ○ |
New, T. R., 2001a. Introduction to the Neuroptera: what are they and how do they operate? In: McEwen, P. K., New, T. R. &
Whittington, A. E. (eds.). Lacewings in the Crop Environment. Cambridge University Press. Cambridge: 3-5.
|
| ○ |
New, T. R., 2001b. Introduction to the systematics and distribution of Coniopterygidae, Hemerobiidae, and Chrysopidae used
in pest management. In: McEwen, P. K., New, T. R. & Whittington, A. E. (eds.). Lacewings in the Crop Environment. Cambridge University Press. Cambridge: 6-28.
|
| ○ |
New, T. R., 2003. The Neuroptera of Malesia. Fauna Malesiana, vol. 4. Brill, Leiden. viii + 204 pp.
|
| ○ |
New, T. R. & Sudarman, H. K., 1988. Neuroptera (Insecta) of the Krakatau Islands, Indonesia. Philosophical Transactions of the Royal Society B: Biological Sciences, 322: 413-426. http://dx.doi.org/10.1098/rstb.1988.0135 |
| ○ |
Newman, E., 1835. Attempted division of British insects into natural orders. Entomological Magazine, 2: 379-431.
|
| ○ |
Newman, E., 1853. Proposed division of Neuroptera into two classes. The Zoologist, 11(Appendix): clxxxi-cciv.
|
| ○ |
Nicoli Aldini, R., 2002. Insetti predatori di interesse agrario. Notiziario sulla Protezione delle Piante, 14: 81-106.
|
| ○ |
Nicoli Aldini, R., Letardi, A. & Pantaleoni, R. A., 2012. State of the art on Neuropterida of Sicily and Malta. Biodiversity Journal, 3(4): 445-458.
|
| ○ |
Nielsen, E. S., 1977. En undersøgelse af netvingefaunaen (Neuroptera s. str.) i en dansk bøgeskov. Entomologiske Meddelelser, 45: 45-64.
|
| ○ |
Nielsen, E. S., 1979. Establishing of synonymy between Helicoconis hirtinervis Tjeder, 1960 and Helicoconis cimbrica Ohm 1965, with description of the larvae (Neuroptera, Coniopterygidae). Nota: Según Meinander (1990: 91) este artículo ha
circulado como manuscrito, pero nunca fue publicado.
|
| ○ |
Obenberger, J., 1958. XXVI. Rad Neuroptera--Hmyz sitokridly. Entomologie, 4: 428-538.
|
| ○ |
Ocharan, R., Ocharan, F. J. & Anadón, A., 2012. Neurópteros de la Reserva de la Biosfera de Muniellos (Asturias, Norte de
España) (Neuroptera s. s., Megaloptera y Raphidioptera). Boletín de la Sociedad entomológica Aragonesa, 50: 421-429.
|
| ○ |
Ohl, M., 2002. Die Neuropterida-Sammlung des Museums für Naturkunde der Humboldt-Universität zu Berlin-Sammler, Forscher,
Typen. Galathea, Supplement 13: 21-29.
|
| ○ |
Ohm, P., 1963. Zur Kenntnis der Coniopterygiden der Kanarischen Inseln. (Neuroptera). Zoologischer Anzeiger, 171,5/8: 264-272.
|
| ○ |
Ohm, P., 1965. Beiträge zur Kenntnis der Gattung Helicoconis Enderlein 1905 (Neuroptera Coniopterygidae) nebst Diagnose zweier neuer Arten aus dem schweizerischen Nationalpark. Ergebnisse der wissenschaftlichen Untersuchungen im schweizerischen Nationalpark, 10: 171-207.
|
| ○ |
Ohm, P., 1968. Vorläufige Beschreibung einer neuen europäischen Aleuropteryx-Art (Neuroptera, Coniopterygidae). Entomologisches Nachrichtenblatt, 15: 12-15.
|
| ○ |
Ohm, P., 1973a. Zwei neue Coniopterygiden-Arten aus Südwesteuropa (Planipennia, Coniopterygidae). Reichenbachia, 14: 237-243.
|
| ○ |
Ohm, P. 1973b. Durch die Forstwirtschaft ermöglichte Vergrösserung der Verbreitungsareale nadelholzbewohnender Netzflügler
(Neuroptera, Planipennia). Faunistisch-Ökologische Mitteilungen, 4:299-304.
|
| ○ |
Ohm, P., 1984. Rote Liste der Netzflügler (Neuroptera). In: J. Blab, E. Nowak, W. Trautmann, H. Sukopp (eds.). Rote liste der gefährdeten Tiere und Pflanzen in der Bundesrepublik Deutschland (4. Auflage. Erweiterte Neubearbeitung). Kilda-Verlag. Steinfurt: 73-75.
|
| ○ |
Ohm, P., 1986. Eine neue Parasemidalis-Art von den Kanarischen Inseln: Parasemidalis similis n.sp. (Planipennia: Coniopterygidae). Entomologische Zeitschrift, 96, 22: 327-331.
|
| ○ |
Ohm, P., 1995. Coniopterygidae in Bernstein-Einschlussen. Eine vorlaufige Ubersicht. Galathea, Supplement 2: 19-20.
|
| ○ |
Ohm, P. & Báez, M., 2004. Planipennia. In: Izquierdo Zamora, Martín Esquivel, Zurita Pérez & Arechavaleta Hernández (eds.).
Lista de especies silvestres de Canarias (Hongos, Plantas y Animales Terrestres). Gobierno de Canarias. La Laguna: 207-208.
|
| ○ |
Ohm, P. & Hölzel, H., 1984. Zur Zoogeographie der Neuropteren auf den Mittelatlantischen Inseln eine erste übersicht (Insecta:
Planipennia). Courier Forschungsinstitut Senckenberg, 71: 79-86.
|
| ○ |
Ohm, P. & Hölzel, H., 1999. Die Coniopterygiden-Gattung Semidalis Enderlein auf den Mittelatlantischen Inseln (Neuropteroidea: Planipennia: Coniopterygidae). Reichenbachia, 33(30): 231-237.
|
| ○ |
Ohm, P. & Remane, R., 1968. Die Neuropterenfauna Hessens und einiger angrenzender Gebiete (Zur Verbreitung der Neuropteren
in Mitteleuropa). Faunistisch-Ökologische Mitteilungen, 3: 209-228.
|
| ○ |
Olivier, G. A., 1792. Encyclopedie méthodique. Histoire naturelle. Vol. 7 (Insectes). Paris. 827 pp.
|
| ○ |
Oswald, J. D., 1993. Revision and cladistic analysis of the world genera of the family Hemerobiidae (Insecta: Neuroptera).
Journal of the New York Entomological Society, 101(2): 143-299.
|
| ○ |
Oswald, J. D., 1996. A new brachypterous Nusalala species from Costa Rica, with coments on the evolution of flightlessness in brown lacewings (Neuroptera: Hemerobiidae). Systematic Entomology, 21(4): 343-352. http://dx.doi.org/10.1111/j.1365-3113.1996.tb00603.x |
| ○ |
Oswald, J. D., 2013a. Neuropterida species of the world. A catalogue of the Species-Group names of the Extant and Fossil Neuroptera,
Megaloptera, Raphidioptera and Glosselytrodea (Insecta: Neuropterida) of the World. Version 3.0. Disponible en: http://lacewing.tamu.edu/Species-Catalogue/index.html (última consulta diciembre de 2015).
|
| ○ |
Oswald, J. D., 2013b. Bibliography of the Neuropterida. An Annotated Bibliography and Digital Library of the Literature of
the Extant and Fossil Neuroptera, Megaloptera, Raphidioptera and Glosselytrodea (Insecta: Neuropterida) of the World, Version
10.0. Disponible en: http://lacewing.tamu.edu/Bibliography/index.html (última consulta diciembre de 2015).
|
| ○ |
Oswald, J. D. & Meinander, M., 1988. Case 2651. Aleuropteryx Löw, 1885 (Insecta, Neuroptera): proposed designation of Aleuropteryx loewii Klapálek, 1894 as the type species. Bulletin of Zoological Nomenclature, 45: 272-274.
|
| ○ |
Oswald, J. D. & Penny, N. D., 1991. Genus-group names of the Neuroptera, Megaloptera and Raphidioptera of the world. Occasional Papers of the California Academy of Sciences, 147: 1-94. http://dx.doi.org/10.5962/bhl.part.3428 |
| ○ |
Packard, A. S., 1883a. On the classification of the Linnaean orders of Orthoptera and Neuroptera. The American Naturalist, 17: 820-829. http://dx.doi.org/10.1086/273433 |
| ○ |
Packard, A. S., 1883b. On the classification of the orders of Orthoptera and Neuroptera. Annals and Magazine of Natural History, (5), 12: 145-154. http://dx.doi.org/10.1080/00222938309459602 |
| ○ |
Pallas, P. S., 1772. Spicilegia zoologica, quibus novae imprimis et obscurae animalium species iconibus, descriptionibus atque commentariis illustrantur,
cura P. S. Pallas. G. A. Lange. Berolini.
|
| ○ |
Panareda Clopés, J. M., 2011. Flora Catalana. Brau Edicions. Figueres. 724 pp.
|
| ○ |
Panareda Clopés, J. M. & Nuet Badia, J., 1993-1994. Tipología y cartografía corológica de las plantas vasculares de Montserrat
(Cordillera Prelitoral Catalana). Revista de Geografía, 27-28: 33-58.
|
| ○ |
Panfilov, D. V., 1980. Novye predstaviteli setcharokrylykh (Neuroptera) iz yury Karatau. In: Dolin, V. G., Panfilov, D. V.,
Ponomarenko, A. G. & Pritykina, L. N. (eds.). Iskopaemye nasekomye mezozoya. Naukova Dumka. Kiev: 82-111.
|
| ○ |
Pantaleoni, R. A., 1983. Neuroptera Planipennia del comprensorio della Valli di Comacchio: indagine ecologica. Bollettino dell’Istituto di Entomologia della Università degli Studi di Bologna, 37: 1-73.
|
| ○ |
Pantaleoni, R. A., 1984. Note su alcuni parassiti (s.l.) di Neurotteri Planipenni con segnalazione del ritrovamento di Acari
foretici su di un Crisopide. Bollettino dell’Istituto di Entomologia “Guido Grandi” della Università degli studi di Bologna, 38: 193-203.
|
| ○ |
Pantaleoni, R. A., 1990. I Neurotteri (Neuropteroidea) della Valle del Bidente-Ronco (Appennino Romagnolo). Bollettino dell’Istituto di Entomologia “Guido Grandi” della Università degli studi di Bologna, 44: 89-142.
|
| ○ |
Pantaleoni, R. A., 1995. Neurotteri (Insecta Neuropteroidea) della Pianura Padana: I parchi urbani e rurali come zone di “rifugio
faunistico”. Quaderni della Stazione di Ecologia del Civico Museo di Storia Naturale di Ferrara, 9: 393-397.
|
| ○ |
Pantaleoni, R. A., 1996. Distribuzione spaziale di alcuni Neurotteri Planipenni su piante arboree. Bollettino dell’Istituto di Entomologia “Guido Grandi” della Università degli studi di Bologna, 50: 133-141.
|
| ○ |
Pantaleoni, R. A., 2007. Perspectivas del uso de Raphidioptera y Neuroptera Coniopterygidae como agentes de control biológico.
In: Rodríguez del Bosque, L. A. & Arredondo Bernal, H. C. (eds.). Teoría y Aplicación del Control Biológico. Sociedad Mexicana de Control Biológico. México D.F.: 106-126.
|
| ○ |
Pantaleoni, R. A., 2010. Nomenclatural roots of Neuropterida: Linnaeus’ era. In: Devetak, D., Lipovšek, S. & Arnett, A. E.
(eds.). Proceedings of the 10th International Symposium on Neuropterology, Piran 2008. University of Maribor. Maribor: 215-229.
|
| ○ |
Pantaleoni, R. A. & Letardi, A., 1996. A remarkable brachypterous female of Dilaridae (Dilar parthenopaeus Costa ?) (Neuroptera). Abstract from the Program of the XX International Congress of Entomology, Firenze: 62.
|
| ○ |
Penny, N. D., Arias, J. R. & Armistead, J. S., 2007. Seasonal emergence of Neuroptera in Fairfax County, Virginia. Proceedings of the California Academy of Sciences, (4), 58: 7-19.
|
| ○ |
Pérez-de la Fuente, R., 2012. Paleobiología de los artrópodos del ámbar cretácico de El Soplao (Cantabria, España). PhD thesis. Universitat de Barcelona. 109 pp.
|
| ○ |
Perkins, R. C. L., 1899. Neuroptera. In: Sharp, D. (ed.). Fauna Hawaiiensis being the land-fauna of the Hawaiian Islands. Vol. 2, pt. 2 (Neuroptera). Cambridge University Press. London: 31-89.
|
| ○ |
Perkins, R. C. L., 1910. Supplement to Neuroptera. In: Sharp, D. (ed.). Fauna Hawaiiensis being the land-fauna of the Hawaiian Islands. Vol. 2. Cambridge University Press. London: 691-696.
|
| ○ |
Perrichot, V., Garrouste, R., Azar, D., Néraudeau, D. & Nel, A., 2014. A new genus of dustywings (Neuroptera: Coniopterygidae)
in Late Cretaceous Vendean amber. Paleontological Contributions, 10F: 25-29.
|
| ○ |
Peters, R. S., Meusemann, K., Petersen, M., Mayer, C., Wilbrandt, J., Ziesmann, T., Donath, A., Kjer, K. M., Aspöck, U., Aspöck,
H., Aberer, A., Stamatakis, A., Friedrich, F., Hünefeld, F., Niehuis, O., Beutel, R. G. & Misof, B., 2014. The evolutionary
history of holometabolous insects inferred from transcriptome-based phylogeny and comprehensive morphological data. BMC Evolutionary Biology, 14: 52. http://dx.doi.org/10.1186/1471-2148-14-52 |
| ○ |
Peterson, A., 1967. Neuroptera. In: Peterson, A. Larvae of Insects, Vol. 2. Coleoptera, Diptera, Neuroptera, Siphonaptera, Mecoptera, Trichoptera. Edwards Brothers. Ann Arbor: 352-361.
|
| ○ |
Pictet, F. J., 1836. Mémoire sur le genre Sialis de Latreille, et considérations sur la classification de l’ordre des Névroptères. Annales des Sciences Naturelles, Zoologie et Biologie Animale, (2), 5: 69-80.
|
| ○ |
Pictet, F. J., 1865. Synopsis des Névroptères d′Espagne. Section II Neuroptera. H.G.Baillière & F.Savy. Genève.
|
| ○ |
Plant, C. W., 1991. An introduction to the British wax-flies (Neuroptera: Coniopterygidae) with a revised key to British species.
British Journal of Entomology and Natural History, 4: 99-117.
|
| ○ |
Plant, C. W., 1992a. A working list of the lacewings of the London Area. London Naturalist, 71: 117-136.
|
| ○ |
Plant, C. W., 1992b. Semidalis pseudouncinata Meinander, 1963 (Neuroptera: Coniopterygidae) new to Britain with a note on its separation from S. aleyrodiformis (Stephens). Entomologist’s Gazette, 43: 292-296.
|
| ○ |
Plant, C. W., 2001. Lacewings (Neuroptera), snakeflies (Raphidioptera), alderflies (Megaloptera) and scorpion flies (Mecoptera)
in Buckingham Palace Garden. In: Plant, C. W. (ed.). The natural history of Buckingham Palace Garden, London. Part 2. London Naturalist 80 (Supp.): 159-167.
|
| ○ |
Poda, N., 1761. Insecta musei Graecensis, quae in ordines, genera et species juxta systema naturae Caroli Linnaei digessit. Jaonnem Baptistam Dietrich. Graecii. 127 + [index] pp. Notas: Edición Facsímil: 1915, W. Junk, Berlin. Neuropterida: “Neuroptera”
(sensu lato), pp. 11-12, 95-101.
|
| ○ |
Poggi, R., 1993. Catalogo dei tipi di Neurotteroidei del Museo Civico di Storia Naturale G.Doria di Genova (Insecta). Annali del Museo Civico di Storia Naturale Giacomo Doria, 89: 415-444.
|
| ○ |
Popov, A., 1973. Über die präimaginalen Stadien paläarktischer Vertreter der Ordnung Neuroptera und Versuch einer neuen systematischen
Gruppierung der Familien mit Rücksicht auf ihre morphologischen und ökologischen Besonderheiten. Bulletin de l’Institut de Zoologie et Musée, Academie Bulgare des Sciences, Section de Biology, 37: 79-101.
|
| ○ |
Popov, A., 1986. Coniopterygiden aus Bulgarien (Neuroptera). Entomologische Nachrichten und Berichte, 30: 167-171.
|
| ○ |
Popov, A., 2002a. Zoogeographical analysis of Neuroptera in Bulgaria. In: Sziráki, G. (ed.). Neuropterology 2000 (Proceedings of the Seventh International Symposium on Neuropterology, Budapest 2000). Acta Zoologica Academiae Scientiarum Hungaricae, 48(Suppl. 2): 271-280.
|
| ○ |
Popov, A., 2002b. Neuropterida of northern Europe. In: Sziráki, G. (ed.). Neuropterology 2000 (Proceedings of the Seventh International Symposium on Neuropterology, Budapest 2000). Acta Zoologica Academiae Scientiarum Hungaricae, 48(Suppl. 2), 281-291 pp.
|
| ○ |
Principi, M. M. & Canard, M., 1974. Les Névroptères. In: Les organismes auxiliaires en verger de pommiers, OILB-SROP. Wageningen: 151-162.
|
| ○ |
Pröse, H., 1992. Rote Liste der gefährdeten Netzflügler (Neuropteroidea) Bayerns. Bayerisches Landesamt für Umweltschutz: Beiträge zum Artenschutz, 15: 137-139.
|
| ○ |
Pröse, H. & Gruppe, A. [V.], 2003. Rote Liste gefährdeter Netzflügler (Neuropteroidea) Bayerns. Schriftenreihe Bayerisches Landesamt für Umweltschutz, 166: 95-98.
|
| ○ |
Putman, W. L. & Herne, D. H. C., 1966. The role of predators and other biotic agents in regulating the population density
of phytophagous mites in Ontario peach orchards. The Canadian Entomologist, 98: 808-820. http://dx.doi.org/10.4039/Ent98808-8 |
| ○ |
Quayle, H. J., 1912. Red spiders and mites of citrus trees. University of California Agricultural Experiment Station Bulletin, 234: 483-530.
|
| ○ |
Quayle, H. J., 1913. Some natural enemies of spiders and mites. Journal of Economic Entomology, 6: 85-88. http://dx.doi.org/10.1093/jee/6.1.85 |
| ○ |
Rambur, J. P., 1842. Histoire Naturelle des Insectes, Névroptères. Librairie encyclopédique de Roret, Fain et Thunot. Paris. [xviii] + 534 pp.
|
| ○ |
Rausch, H. & Aspöck, H., 1977. Coniopteryx (Holoconiopteryx) renate n. sp. (Neuroptera, Planipennia). Zeitschrift der Arbeitsgemeinschaft Österreichischer Entomologen, 29: 72.
|
| ○ |
Rausch, H. & Aspöck, H., 1978a. Zwei neue Spezies des Genus Aleuropteryx Löw aus dem westlichen Mittelmeergebiet (Neuroptera, Coniopterygidae). Nachrichtenblatt der Bayerischen Entomologen, 27: 9-13.
|
| ○ |
Rausch, H. & Aspöck, H., 1978b. Zwei neue Spezies des Genus Nimboa Navás (Neuroptera, Coniopterygidae) aus Vorderasien. Zeitschrift der Arbeitsgemeinschaft Österreichischer Entomologen, 30: 13-16.
|
| ○ |
Rausch, H. & Aspöck, H., 1978c. Drei neue Spezies des Genus Coniopteryx Curtis (Neuroptera, Coniopterygidae) aus dem Iran. Zeitschrift der Arbeitsgemeinschaft Österreichischer Entomologen, 29: 100-104.
|
| ○ |
Rausch, H., Aspöck, H. & Aspöck, U., 1978a. Beschreibung von Helicoconis sengonca n. sp., einer neuen Coniopterygiden-Spezies aus Anatolien, und Bemerkungen über Helicoconis aptera Messner, 1965 (Neuropteroidea, Planipennia). Zeitschrift der Arbeitsgemeinschaft Österreichischer Entomologen, 30: 25-28.
|
| ○ |
Rausch, H., Aspöck, H. & Aspöck, U., 1981. Helicoconis tjederi n. sp. und Helicoconis premnata n. sp.--zwei neue Coniopterygiden-Spezies aus dem West-Himalaya (Neuropteroidea, Planipennia). Nachrichtenblatt der Bayerischen Entomologen, 30: 8-12.
|
| ○ |
Rausch, H., Aspöck, H. & Ohm, P., 1978b. Zwei weitere neue Arten des Genus Aleuropteryx aus der Westpaläarktis (Neuroptera, Coniopterygidae). Entomologische Zeitschrift, 88: 45-49.
|
| ○ |
Ren, D. & Hong, Y.-c., 1994. A cladistic study on the familial phylogeny of fossil and living Raphidioptera (Insecta). Bulletin of the Chinese Academy of Geological Sciences, 29: 103-117.
|
| ○ |
Ressl, F., 1964a. Zur Erforschung der Wärmeinsel im Heidegebiet Schauboden-Hochriess. Entomologisches Nachrichtenblatt, 11: 9-12.
|
| ○ |
Ressl, F., 1964b. Abundanzverhältnisse der Neuropteren des Verwaltungsbezirkes Scheibbs, N.O. Entomologisches Nachrichtenblatt, 11: 63-68.
|
| ○ |
Ressl, F., 1971. Untersuchungen über die Coniopterygiden (Neuroptera, Planipennia) des Bezirkes Scheibbs (NÖ)-Ein Beitrag
zur Kenntnis der Verbreitung, Phänologie und Ökologie der Coniopterygiden Mitteleuropas. Nachrichtenblatt der Bayerischen Entomologen, 20: 44-60.
|
| ○ |
Reuter, O. M., 1894. Neuroptera Fennica. Förteckning och Beskrifning öfver Finlands Neuropterer. Acta Societatis pro Fauna et Flora Fennica, 9(8): 1-36.
|
| ○ |
Ribeiro, F. N., 1997. Protecção integrada em pomares de macieiras na rigiao de Alcobaça 4. Contribução para o conhecimento
da fauna útil em Quercus coccifera L. Boletim da Sociedade Portuguesa de Entomologia, 169(VI-19): 253-264.
|
| ○ |
Ribera, I. & Blasco-Zumeta, J., 1998. Biogeographical links between steppe insects in the Monegros region (Aragón, NE Spain),
the eastern Mediterranean, and central Asia. Journal of Biogeography, 25: 969-986. http://dx.doi.org/10.1046/j.1365-2699.1998.00226.x |
| ○ |
Riek, E. F., 1967. Structures of unknown, possibly stridulatory, function of the wings and body of Neuroptera; with an appendix
on other endopterygote orders. Australian Journal of Zoology, 15: 337-348.
|
| ○ |
Riek, E. F., 1970. Neuroptera (Lacewings). In: The insects of Australia. Melbourne University Press. Melbourne: 472-494.
|
| ○ |
Riek, E. F., 1975. On the phylogenetic position of Brucheiser argentinus Navás 1927 and description of a second species from Chile (Insecta: Neuroptera). Studies on the Neotropical Fauna, 10:117-126. http://dx.doi.org/10.1080/01650527509360487 |
| ○ |
Ripolles, J.L. & Melia, A., 1980. Primeras observaciones sobre la proliferación de Conwentzia psociformis (Curt.) (Neuroptera, Coniopterygidae), en los cítricos de Castellón de la Plana. Boletín de Sanidad Vegetal, Plagas, 14: 5-9.
|
| ○ |
Roepke, W., 1916. Eine neue Coniopterygidae aus Java (Parasemidalis decipiens n. g. n. sp.). Zoologische Mededeelingen, 2: 156-158.
|
| ○ |
Roepke, W., 1917. Das Gespinst von Parasemidalis (Neuroptera, Coniopterygidae). Zoologische Mededeelingen, 3: 274.
|
| ○ |
Röhricht, W., 2002. Neuroptera.com. Neuropteren im Internet-nutzt das was? Galathea, Supplement 13: 31-36.
|
| ○ |
Röhricht, W., 2004. Rote Liste der Netzflügler i. w. S. (Neuropterida) des Landes Sachsen-Anhalt (2. Fassung, Stand: Februar
2004). Berichte des Landesamtes für Umweltschutz Sachsen-Anhalt, 39: 249-251.
|
| ○ |
Röhricht, W. & Tröger, E. J., 1998. Rote Liste der Netzflügler (Neuropteroidea). In: Binot, M., Bless, R., Boye, P., Gruttke,
H. & Pretscher, P. (eds.). Rote Liste gefährdeter Tiere Deutschlands. Schriftenreihe für Landschaftspflege und Naturschutz, 55: 231-234.
|
| ○ |
Ros, J., Moner, P., Roig, V., Castillo, E. & Lorite, P., 1988. Eficacia del hidrolizado de proteína en las pulverizaciones-cebo
contra Ceratitis capitata Wied. Boletín de Sanidad Vegetal, Plagas, 14: 5-9.
|
| ○ |
Rossi, P., 1790. Fauna Etrusca sistens insecta quae in provinciis Florentina et Pisana praesertim collegit Petrus Rossius. Vol. 2. Th. Masi & Sociorum. Liburni. 348 pp.
|
| ○ |
Rostock, M. & Kolbe, H., 1888. Neuroptera germanica. Die Netzflügler Deutschlands mit Berücksichtigung auch einiger ausserdeutscher
Arten nach der analytischen Methode unter Mitwirkung von H. Kolbe bearbeitet. Jahresbericht des Vereins für Naturkunde zu Zwickau, 1887: 1-198.
|
| ○ |
Rousset, A., 1956a. Sur l’anatomie céphalique des larves de Coniopterygidae (Névroptères Planipennes). Comptes Rendus Hebdomadaires des Séances de l’Académie des Sciences, 242: 934-936.
|
| ○ |
Rousset, A., 1956b. Sur l’anatomie céphalique des larves de Coniopterygidae (Névroptères Planipennes). Les stylets et leur
musculature. Comptes Rendus Hebdomadaires des Séances de l’Académie des Sciences, 243: 869-872.
|
| ○ |
Rousset, A., 1958. Sur le système nerveux central céphalique de la larve de Coniopteryx (Névropt. Planipenne). Comptes Rendus Hebdomadaires des Séances de l’Académie des Sciences, 246: 842-845.
|
| ○ |
Rousset, A., 1960a. Etude statistique d’une population de Coniopteryx pygmaea End. (Nevropteres Planipennes). Travaux du Laboratoire de Zoologie et de la Station Aquicole Grimaldi de la Faculté des Sciences de Dijon, 34: 1-20.
|
| ○ |
Rousset, A., 1960b. Contribution à la faune de France des Névroptères. Travaux du Laboratoire de Zoologie et de la Station Aquicole Grimaldi de la Faculté des Sciences de Dijon, 35: 23-33.
|
| ○ |
Rousset, A., 1964. Description d’une espèce nouvelle du genre Coniopteryx Curt., Coniopteryx drammonti (Névroptères Planipennes Conioptérygidés). Travaux du Laboratoire de Zoologie et de la Station Aquicole Grimaldi de la Faculté des Sciences de Dijon, 56: 1-10.
|
| ○ |
Rousset, A., 1966. Morphologie céphalique des larves de planipennes (Insectes Névroptéroïdes). Mémoires du Museum Nationale d’Histoire Naturelle, Série A Zoologie, 42: 1-199.
|
| ○ |
Rousset, A., 1968. Déplacements d’attaches musculaires et formation de structures endosquelettiques chez les insectes: cas
de la musculature spinale des larves de Conioptérygidés (Névroptères, Planipennes). Comptes Rendus Hebdomadaires des Séances de l’Académie des Sciences, D, Sciences Naturelles, 267: 1458-1461.
|
| ○ |
Rousset, A., 1969. Morphologie thoracique des larves de Planipennes (Insectes Névroptéroîdes). I. Squlette et musculature
des régions antérieures du thorax chez les larves de Chrysopa et de Conioptérygidés. Annales des Sciences Naturelles, Zoologie et Biologie Animale, (12), 11: 97-138.
|
| ○ |
Rousset, A., 1970. Morphologie thoracique des larves de Planipennes (Insectes Névroptéroîdes). II. Innervation motrice thoracique
et trajet des fribres dans les systèmes des nerfs intercalaires. Annales des Sciences Naturelles, Zoologie et Biologie Animale, (12), 12: 479-503.
|
| ○ |
Ruiz, M. & Montiel, A., 2001. Introduccción al conocimiento de la entomofauna del olivar en la provincia de Jaén. Aspectos
cuantitativos (II). Boletín de Sanidad Vegetal, Plagas, 27: 531-560.
|
| ○ |
Saure, C. & Gerstberger, M., 1991. Standardliste und Rote Liste der Neuropteroidea (Netzflügler s.l.) von Berlin. In: Auhagen,
A., Platen, R. & Sukopp, H. (eds.). Rote listen der gefährdeten Pflanzen und Tiere in Berlin. Landschaftsentwicklung und Umweltforschung, 6. Sonderheft: 237-241.
|
| ○ |
Schedl, W., 1970. Planipennia (Insecta, Neuroptera) der subalpinen und höheren Stufen der Ötztaler Alpen (Tirol, Österreich).
Bericht des Naturwissenschaftliche-Medizinischen Vereins in Innsbruck, 58: 305-312.
|
| ○ |
Schlechtendal, D. H. R., von., 1882. Coniopteryx psociformis Curtis, als Schmarotzer in Spinneneiern. Jahresbericht des Vereins für Naturkunde zu Zwickau, 1881: 26-31.
|
| ○ |
Schlechtendal, D. H. R., von., 1883. Nachträgliche Berichtigung über Coniopteryx psociformis Curtis. Jahresbericht des Vereins für Naturkunde zu Zwickau, 1882: 70-72.
|
| ○ |
Schlüter, T., 1986. The fossil Planipennia-a review. In: Gepp, J., Aspöck, H. & Hölzel, H. (Eds.). Recent Research in Neuropterology (Proceedings of the 2nd International Symposium on Neuropterology, Hamburg 1984). Graz: 103-111.
|
| ○ |
Schrank von Paula, F., 1781. Envmeratio insectorvm Avstriae indigenorum. Klett & Franck. Avgvstae Vindelicorvm. [xxii] + 548 pp. 4 plates.
|
| ○ |
Schremmer, F., 1959. Beobachtungen und Untersuchungen über die Insektenfauna der Lärche (Larix decidua) im östlichen Randgebiet ihrer natürlichen Verbreitung, mit besonderer Berücksichtigung einer Grossstadtlärche. Teil I u.
II. Zeitschrift für Angewandte Entomologie, 45: 1-48, 113-153.
|
| ○ |
Scopoli, J. A., 1763. Entomologia Carniolica, exhibens insecta Carnioliae indigena et distributa in ordines, genera, species, varietates, methodo
Linneana. Ioannis Thomae Trattner. Vindobonae. [38] + 418 + [1] pp.
|
| ○ |
Séméria, Y. & Berland, L., 1988. Atlas des Névroptères de France et d’Europe. Mégaloptères, Raphidioptères, Nèvroptères Planipennes, Mécoptères. Nouvelle [2ème] édition revue et augmentée. Boubée. Paris. 190 pp.
|
| ○ |
Silvestri, F., 1942. Ordo Neuroptera. In: Compendio di Entomologia Applicata (Agraria-Forestale-Medica-Veterinaria), Vol. 2. E. della Torre. Portici: 1-33.
|
| ○ |
Simpson, E.M., 1949. Measurement of diversity. Nature, 163: 688. http://dx.doi.org/10.1038/163688a0 |
| ○ |
Sinacori, A., Mineo, G. & Lo Verde, G., 1992. Osservazioni su Aphanogmus steinitzi Priesner (Hym. Ceraphronidae) parassitoide di Conwentzia psociformis (Curtis) (Neur. Coniopterygidae). Phytophaga, 4: 29-48.
|
| ○ |
Smith, R. C., 1934. Notes on the Neuroptera and Mecoptera of Kansas, with keys for the identification of species. Journal of the Kansas Entomological Society, 7: 120-145.
|
| ○ |
Soler, J. M., García Marí, F. & Alonso, D., 2002. Evolución estacional de la entomofauna auxiliar en cítricos. Boletín de Sanidad Vegetal, Plagas, 28: 133-149.
|
| ○ |
Southwood, T.R.E., 1957. Observations on swarming in Braconidae (Hymenoptera) and Coniopterygidae (Neuroptera). Proceedings of the Royal Entomological Society of London. Series A, General Entomology, 32: 80-82. http://dx.doi.org/10.1111/j.1365-3032.1957.tb00376.x |
| ○ |
Stange, L. A., 1981. The Dustywings of Florida. Part. 1. Genera (Neuroptera: Coniopterygidae). Florida Department of Agriculture and Consumer Service, Entomology Circular, 233: 1-2.
|
| ○ |
Stange, L. A. & Wang, H.-y., 1998. Guide book to insects in Taiwan. 18. Neuroptera, Megaloptera, Raphidioptera. Hsu Hsin Books. Taipei. viii + 278 pp.
|
| ○ |
Starý, B., Bezděčka, P., Čapek, M., Starý, P., Zelený, J. & Šedivý, J., 1988. Atlas of insects beneficial to forest trees, Volume 2. Elsevier. Amsterdam. 100 pp.
|
| ○ |
Stelzl, M., 1989. Nahrungsspektren mitteleuropäischer Neuropteren-Imagines (Neuropteroidea: Insecta). Diplomarbeit. Universität Graz. 145 pp.
|
| ○ |
Stelzl, M., 1991. Untersuchungen zu Nahrungsspektren mitteleuropäischer Neuropteren-Imagines (Neuropteroidea, Insecta) Mit
einer Diskussion über deren Nützlichkeit als Opponenten von Pflanzenschädlingen. Journal of Applied Entomology, 111: 469-477. http://dx.doi.org/10.1111/j.1439-0418.1991.tb00349.x |
| ○ |
Stelzl, M., 1992. Comparative studies on mouthparts and feeding habits of adult Raphidioptera and Neuroptera (Insecta: Neuropteroidea).
In: Canard, M., Aspöck, H. & Mansell, M. W. (eds.). Current Research in Neuropterology (Proceedings of the Fourth International Symposium on Neuropterology, Bagnères-de-Luchon
1991). Toulouse: 341-347.
|
| ○ |
Stelzl, M. & Devetak, D., 1999. Neuroptera in agricultural ecosystems. Agriculture, Ecosystems & Environment, 74: 305-321. http://dx.doi.org/10.1016/S0167-8809(99)00040-7
|
| ○ |
Stephens, J. F., 1836. Illustrations of British entomology; or, a synopsis of indigenous insects: containing their generic and specific distinctions;
with an account of their metamorphoses, times of appearance, localities, food, economy, as far as practicable. Mandibulata. Vol. 6. Baldwin and Cradock, London. 240 pp.
|
| ○ |
Stimmel, J. F., 1979. Seasonal history and distribution of Carulaspis minima (Targ.-Tozz.) in Pennsylvania (Homoptera: Diaspididae). Proceedings of the Entomological Society of Washington, 81: 222-229.
|
| ○ |
Stitz, H., 1927. 17.-20. Ordnung: Netzflügler, Neuroptera. In: P. Brohmer, P. Ehrmann & G. Ulmer (eds.). Die Tierwelt Mitteleuropas, 6(14). Quelle and Meyer. Leipzig: 1-23.
|
| ○ |
Stitz, H., 1928. Die Neuropteren des arktischen Gebietes. Fauna Arctica, 2(5): 634-638, 955.
|
| ○ |
Stitz, H., 1931. Planipennia. In: Schultze, P. (ed.). Biologie der Tiere Deutschlands, 33(35). Borntraeger. Berlin: 67-304.
|
| ○ |
Sugg, P. M., Greve, L. & Edwards, J. S., 1994. Neuropteroidea from Mount St. Helens and Mount Rainier: dispersal and immigration
in volcanic landscapes. Pan-Pacific Entomologist, 70: 212-221.
|
| ○ |
Sziráki, G., 1979. Notes on Hungarian species of Coniopterygidae from different orchards. Folia Entomologica Hungarica (N.S.), 32: 181-184.
|
| ○ |
Sziráki, G., 1989. A magyar faunára új két lisztesfátyolka (Planipennia: Coniopterygidae). Folia Entomologica Hungarica (N.S.), 50: 185-186.
|
| ○ |
Sziráki, G., 1990. Two Aleuropteryx species from Tanzania (Planipennia: Coniopterygidae). Rovartani Közlemények (N.S.), 51: 117-121.
|
| ○ |
Sziráki, G., 1991. A new Coniopteryx species from the far east (Planipennia: Coniopterygidae). Folia Entomologica Hungarica (N.S.), 52: 109-111.
|
| ○ |
Sziráki, G., 1992a. Coniopterygidae of Hungary with a key to the identification of Coniopteryx Curtis females (Insecta: Neuroptera:
Coniopterygidae). In: Canard, M., Aspöck, H. & Mansell, M. W. (eds.). Current Research in Neuropterology (Proceedings of the Fourth International Symposium on Neuropterology, Bagnères-de-Luchon
1991). Toulouse: 359-366.
|
| ○ |
Sziráki, G., 1992b. A possibility for the identification of female coniopterygids (Neuroptera). Proceedings of the 4th ECE/XIII. SIEEC, Gödöllö 1991: 110-114.
|
| ○ |
Sziráki, G., 1992c. Female internal genitalia of the Coniopteryx species of central Europe (Neuroptera, Coniopterygidae). Acta Zoologica Hungarica, 38: 359-371.
|
| ○ |
Sziráki, G., 1992d. A new Coniopteryx species from India (Neuroptera: Coniopterygidae). Folia Entomologica Hungarica (N.S.), 53: 189-191.
|
| ○ |
Sziráki, G., 1992e. Coniopterygidae from Yemen (Neuroptera). Acta Zoologica Hungarica, 38: 89-94.
|
| ○ |
Sziráki, G., 1994a. Coniopterygidae from the Afrotropical Region (Neuroptera). Acta Zoologica Academiae Scientiarum Hungaricae, 40: 171-184.
|
| ○ |
Sziráki, G., 1994b. Helicoconis transsylvanica Kis, 1965 (Neuroptera: Coniopterygidae) magyarországi elöfordulása a faj hím genitáliájának ismeretére vonatkozó megjegyzésekkel.
Folia Entomologica Hungarica, (N.S.), 55: 408-409.
|
| ○ |
Sziráki, G., 1996. The internal genitalia of females of some coniopterygid genera, compared with other neuropteroid taxa (Insecta:
Neuroptera: Coniopterygidae). In: Canard, M., Aspöck, H. & Mansell, M. W. (eds.). Current Research in Neuropterology (Proceedings of the Fourth International Symposium on Neuropterology, Bagnères-de-Luchon
1991). Toulouse: 217-228.
|
| ○ |
Sziráki, G., 1997. Data to the coniopterygid fauna of Yemen, with description of twelve new species (Neuroptera: Coniopterygidae).
Acta Zoologica Academiae Scientiarum Hungaricae, 43: 271-294.
|
| ○ |
Sziráki, G., 1998a. Female internal genitalia of some Neuroptera of phylogenetic interest. In: Panelius, S. P. (ed.). Neuropterology 1997 (Proceedings of the Sixth International Symposium on Neuropterology, Helsinki 1997). Acta Zoologica Fennica, 209: 243-247.
|
| ○ |
Sziráki, G., 1998b. Zoogeographic relations of South Asian coniopterygids (Neuroptera, Coniopterygidae). In: Panelius, S.
P. (ed.). Neuropterology 1997 (Proceedings of the Sixth International Symposium on Neuropterology, Helsinki 1997). Acta Zoologica Fennica, 209: 249-254.
|
| ○ |
Sziráki, G., 1999. Coniopterygidae (Neuroptera) from southwestern parts of Asia. Folia Entomologica Hungarica (N.S.), 60: 179-186.
|
| ○ |
Sziráki, G., 2001. Coniopterygidae from Vietnam (Neuroptera). Rovartani Közlemények (N.S.), 62: 51-63.
|
| ○ |
Sziráki, G., 2002a. Contribution to knowledge of female internal genitalia of Neuroptera. In: Sziráki, G. (ed.). Neuropterology 2000 (Proceedings of the Seventh International Symposium on Neuropterology, Budapest 2000). Acta Zoologica Academiae Scientiarum Hungaricae, 48(Suppl. 2): 341-349.
|
| ○ |
Sziráki, G., 2002b. Coniopterygidae (Neuroptera) from Thailand. Rovartani Közlemények (N.S.), 63: 53-64.
|
| ○ |
Sziráki, G., 2004. Coniopterygidae of Eurasia, New Guinea and North Africa. Folia Entomologica Hungarica (N.S.), 65(Suppl.): 1-166.
|
| ○ |
Sziráki, G., 2005. Species grouping of several genera and subgenera of Coniopterygidae (Neuroptera). Folia Entomologica Hungarica (N.S.), 66: 101-136.
|
| ○ |
Sziráki, G., 2007a. Studies on Brucheiserinae (Neuroptera: Coniopterygidae), with description of the second genus of the subfamily.
Acta Zoologica Academiae Scientiarum Hungaricae, 53(Suppl. 1): 231-254.
|
| ○ |
Sziráki, G., 2007b. Presence of the subgenus Xeroconiopteryx Meinander, 1972 in Hungary (Neuroptera: Coniopterygidae). Annales Historico-Naturales Musei Nationalis Hungarici, 99: 93-100.
|
| ○ |
Sziráki, G., 2009. Data on Coniopterygidae of Argentina, with subgeneric division of Parasemidalis Enderlein, 1905. Folia Historico-Naturalia Musei Matraensis, 33: 169-199.
|
| ○ |
Sziráki, G., 2010a. Taxonomic and nomenclatural questions of some coniopterygid species (Neuroptera). Folia Historico-Naturalia Musei Matraensis, 34: 141-150.
|
| ○ |
Sziráki, G., 2010b. Order Neuroptera, family Coniopterygidae. In: van Harten, A. (ed.). Arthropod fauna of the United Arab Emirates, 3. Dar Al Ummah. Abu Dhabi: 283-298.
|
| ○ |
Sziráki, G., 2010c. Taxonomic and nomenclatural questions of some coniopterygid species (Neuroptera). Folia Historico-Naturalia Musei Matraensis, 34: 141-150.
|
| ○ |
Sziráki, G., 2011. Coniopterygidae of the world. Annotated check-list and identification keys for living species, species groups and supraspecific
taxa of the family. Lambert Academic Publishing. Saarbrücken. 249 pp. (nos ha sido imposible su consulta o su adquisición por encontrarse agotada
la edición de la obra).
|
| ○ |
Sziráki, G., 2015. Identity of Coniopteryx madagascariensis Meinander, 1974 (Neuroptera: Coniopterygidae), with description of three new species. Acta Zoologica Academiae Scientiarum Hungaricae, 61(2): 135-146. http://dx.doi.org/10.17109/AZH.61.2.135.2015 |
| ○ |
Sziráki, G. & Flint, O. S., 2007. Larva of Brucheiser penai Riek, 1975 (Neuroptera Coniopterygidae). In: R. A. Pantaleoni, A. Letardi & C. Corazza (eds), Proceedings of the IX International Symposium on Neuropterology, Ferrara 2005. Annali del Museo civico di Storia naturale di Ferrara, 8: 45-48.
|
| ○ |
Sziráki, G. & Greve, L., 1996. Some Coniopterygidae (Neuroptera) from a mountain rainforest of Tanzania. Acta Zoologica Academiae Scientiarum Hungaricae, 42(1): 81-88.
|
| ○ |
Sziráki, G. & Greve, L., 2000. Parasemidalis principiae sp. n.--the first Parasemidalis species from South America (Neuroptera: Coniopterygidae). Journal of Neuropterology, 3: 5-8.
|
| ○ |
Sziráki, G. & Harten, A. van, 2006. The dusty lacewings (Neuroptera, Coniopterygidae) of the Arabian Peninsula. Fauna of Arabia, 22: 381-434.
|
| ○ |
Sziráki, G. & Mirmoayedi, A., 2012. Annotated checklist of the Iranian Coniopterygidae (Neuroptera). Folia Historico-Naturalia Musei Matraensis, 36: 45-50.
|
| ○ |
Sziráki, G. & Penny, N. D., 2012. Data on the dusty lacewing fauna of northwestern Argentina with description of a new species
(Neuroptera: Coniopterygidae). Psyche. ID508726: 1-4. http://dx.doi.org/10.1155/2012/508726 |
| ○ |
Sziráki, G. & Winterton, S., 2012. An unusual species of Cryptoscenea Enderlein, 1914 from Western Australia (Neuroptera: Coniopterygidae). Genus, 23: 419-424.
|
| ○ |
Tauber, C. A., 1991. Order Neuroptera. In: Stehr, F. W. (ed.). Immature insects, Vol. 2. Kendall/Hunt Publishing. Dubuque: 126-143.
|
| ○ |
Tauber, C. A. & Adams, P. A., 1990. Systematics of the Neuropteroidea: present status and future needs. In: Kosztarab, M.
& Schaefer, C. W. (eds.). Systematics of the North American Insects and Arachnids: Status and Needs. Virginia Polytechnic Institute and State University. Blacksburg: 151-164.
|
| ○ |
Tauber, C. A., Tauber, M. J. & Albuquerque, G. S., 2009. Neuroptera (lacewings, antlions). In: Resh, V. H. & Cardé, R. T.
(eds.). Encyclopedia of Insects. 2nd Edition. Elsevier. Amsterdam: 695-707.
|
| ○ |
Tauber, C. A., Tauber, M. J. & Giffin, J. G., 2007. Flightless Hawaiian Hemerobiidae (Neuroptera): comparative morphology
and biology of a brachypterous species, its macropterous relative and intermediate forms. European Journal of Entomology, 104: 787-800. http://dx.doi.org/10.14411/eje.2007.100 |
| ○ |
Tellería, J. L., 2013. Pérdida de biodiversidad. Causas y consecuencias de la desaparición de las especies Loss of biodiversity:
causes and consequences of the species loss. Memorias de la Real Sociedad Española de Historia Natural, 2a época, 10: 13-25.
|
| ○ |
Thaler, K., 1997. In memoriam em. Univ.-Prof. Dr. Heinz Janetschek (1913-1997). Berichte des naturwissenschaftlich-medizinischen Vereins in Innsbruck, 84: 411-417.
|
| ○ |
Tillier, P., 2013. Deux espèces du genre Helicoconis Enderlein, 1905, nouvelles pour la France et liste actualisée des Coniopterygidae de France (Neuroptera). Bulletin de la Société entomologique de France, 118(2): 141-144.
|
| ○ |
Tillier, P., 2015. Raphidioptera et Neuroptera (Insecta, Neuropterida) du Parc national du Mercantour (France). In: Daugeron
C., Deharveng L., Isaia M., Villemant C. & Judson M. (eds.). Mercantour/Alpi Marittime All Taxa Biodiversity Inventory. Zoosystema, 37(4): 581-594. http://dx.doi.org/10.5252/z2015n4a4 |
| ○ |
Tillyard, R. J., 1916. Studies in Australian Neuroptera. No. iv. The families Ithonidae, Hemerobiidae, Sisyridae, Berothidae,
and the new family Trichomatidae; with a discussion of their characters and relationships, and descriptions of new and little-known
genera and species. Proceedings of the Linnean Society of New South Wales, 41: 269-332.
|
| ○ |
Tillyard, R. J., 1919. The panorpoid complex. A study of the phylogeny of the holometabolous insects, with special reference
to the subclasses Panorpoidea and Neuropteroidea. Introduction. Proceedings of the Linnean Society of New South Wales, 43: 265-284.
|
| ○ |
Tillyard, R. J., 1926. The Insects of Australia and New Zealand. Angus and Robertson. Sydney. xi + 560 pp.
|
| ○ |
Tillyard, R. J., 1932. Kansas Permian Insects. Part 14. The order Neuroptera. American Journal of Science, (5), 23: 1-30. http://dx.doi.org/10.2475/ajs.s5-23.133.1 |
| ○ |
Tjeder, B., 1930. Preliminary descriptions of three new Coniopteryx-species from northern Europe. Entomologisk Tidskrift, 51: 206.
|
| ○ |
Tjeder, B., 1931. A revision of the north-European species of the genus Coniopteryx Curt. (s. str.), based upon a study of the male and female genitalia. Arkiv för Zoologi, 23A(10): 1-32.
|
| ○ |
Tjeder, B., 1940. Revisions of the Scandinavian Neuroptera (s. str.) and Mecoptera recorded by J. W. Zetterstedt, H. D. J.
Wallengren, and others. Opuscula Entomologica, 5: 67-114.
|
| ○ |
Tjeder, B., 1943. The Neuroptera and Mecoptera of northern Norway (Nordland, Troms and Finnmark). Tromso Museums Årshefter, 63(3): 1-15.
|
| ○ |
Tjeder, B., 1952. Specimina typorum insectorum a Carolo Linnaeo descriptorum in Museo Degeeriano asservata. Pars III. Neuroptera. Entomologisk Tidskrift, 73: 203-206.
|
| ○ |
Tjeder, B., 1954. Genital structures and terminology in the order Neuroptera. Entomologiske Meddelelser, 27: 23-40.
|
| ○ |
Tjeder, B., 1956. Neuroptera. In: Tuxen, S. L. [P.] (ed.). Taxonomist’s glossary of genitalia in insects. Munksgaard. Copenhagen: 76-83.
|
| ○ |
Tjeder, B., 1957. Neuroptera-Planipennia. The Lace-wings of Southern Africa. 1. Introduction and Families Coniopterygidae,
Sisyridae, and Osmylidae. In: Hanström, B., Brinck, P. & Rudebec, G., South African Animal Life. Vol. 6. Swedish Natural Science Research Council. Stockholm: 95-188.
|
| ○ |
Tjeder, B., 1960. A new subapterous coniopterygid from Denmark (Neuroptera). Entomologiske Meddelelser, 29: 312-319.
|
| ○ |
Tjeder, B., 1964. The female of Coniopteryx tullgreni Tj. (Neuroptera, Coniopterygidae). Opuscula Entomologica, 29: 168-169.
|
| ○ |
Tjeder, B., 1967. Kullabergs nätvingar, näbbsländor, nattsländor och harkrankar. Kullabergs Natur, 12: 1-21.
|
| ○ |
Tjeder, B., 1969. New Coniopterygidae from Southern Africa (Neuroptera). Opuscula Entomologica, 34: 243-249.
|
| ○ |
Tjeder, B., 1970. Neuroptera. In: Tuxen, S. L. [P.] (ed.). Taxonomist’s glossary of genitalia in insects. 2nd edition. Munksgaard. Copenhagen: 89-99.
|
| ○ |
Tjeder, B., 1972. Coniopterygidae from the Snow Mountains, New Guinea (Neuroptera). Entomologisk Tidskrift, 93: 186-209.
|
| ○ |
Tjeder, B., 1987. A new species of Coniopteryx from the Namib dunes, S.W.Africa/Namibia (Neuroptera: Coniopterygidae). Neuroptera International, 4(4): 275-277.
|
| ○ |
Tröger, E. J., 1993. Vorläufige Rote Liste gefährdeter Netzflügler (Neuropteroides) in Baden-Wüttemberg (Stand Juli 1992).
In: AAVV, Rote Listen der gefährdeter Tiere und Pflanzen Baden-Wüttembergs, IIIB: 27-28.
|
| ○ |
Tullgren, A., 1906. Zur Kenntnis schwedischer Coniopterygiden. Arkiv för Zoologi, 3(5): 1-15.
|
| ○ |
Tuxen, S. L., [P.] 1970. Taxonomist’s glossary of genitalia in insects. 2nd Edition. Munksgaard. Copenhagen. 359 pp.
|
| ○ |
Valencia, V. L., 2009. Entomología urbana a la memoria de Don Raul Cortés Pena (Q.E.P.D.), mi maestro y amigo. Enemigos naturales
de Aleurodicus juleikae Bondar (Hemiptera: Aleyrodidae) en un ambiente urbano de Lima, Perú. Idesia, 27: 79-89.
|
| ○ |
Vas, J., Ábrahám, L. & Markó, V., 1999. Study of nocturnal and diurnal activities of lacewings (Neuropteroidea: Raphidioptera,
Neuroptera) by suction trap. Acta Phytopathologica et Entomologica Hungarica, 34: 149-152.
|
| ○ |
Vas, J., Ábrahám, L. & Markó, V., 2001. Methodological investigations on a Neuropteroidea community. Acta Phytopathologica et Entomologica Hungarica, 36: 101-113.
|
| ○ |
Vidal y López, M., 1943. Catálogo provisional de Neurópteros de la región levantina. Graellsia, 1(6): 13-24.
|
| ○ |
Vidlička, L., 2014. Description of Helicoconis tatricus sp. n. (Neuroptera, Coniopterygidae) from Slovakia (Central Europe) and key for determination of all Palaearctic species of subgenus Helicoconis. Zootaxa, 3893(3): 438-444. http://dx.doi.org/10.11646/zootaxa.3893.3.8 |
| ○ |
Vidlička, L., 2015. Sieťokrídlovce (Neuroptera) Tatier. Ústav Zoológie Slovenskej Akadémie Vied Scientica. Bratislava. 109 pp.
|
| ○ |
Viggiani, D. G., 1967. Ricerche sugli Hymenoptera Chalcidoidea. XIII. Sugli entomoparassiti della Semidalis aleurodiformis Steph. (Neur. Coniopterygidae), con descrizione di un nuovo genere di Encyrtidae. Bollettino del Laboratorio di Entomologia Agraria “Filippo Silvestri” di Portici, 25: 163-175.
|
| ○ |
Viggiani, G., 1977. Neuroptera. In: Viggiani, G. Lotta Biologica ed Integrata. Liguori Editori. Napoli: 46-56.
|
| ○ |
Vine, H. C. A., 1895. Predacious and parasitic enemies of aphides (including a study of hyper-parasites). Part II. Journal of Microscopy and Natural Science, 14: 33-42, 121-134, 254-268.
|
| ○ |
Walker, F., 1853. List of the specimens of neuropterous insects in the collection of the British Museum. Part II. (Sialides-Nemopterides). British Museum. London. [iii] + 193-476.
|
| ○ |
Wallengren, H. D. J., 1863. Bidrag till kännedomen af Sveriges Neuroptera. Öfversigt af Kongliga Vetenskaps-Akademiens Forhändlingar, 20: 15-26.
|
| ○ |
Wallengren, H. D. J., 1871. Skandinaviens Neuroptera. Kungliga Svenska Wetenskaps Academiens Handlingar (NF), 9(8): 1-76.
|
| ○ |
Wang, W.-q.; Liu, Z.-q., 2007. Molecular phylogenetic relationship of the Coniopterygidae from China based on 16S rRNA sequences.
Acta Zootaxonomica Sinica, 32: 851-855.
|
| ○ |
Wang, W.-q., Ma, L.-l., Sziráki, G. & Liu, Z.-q., 2007. Taxonomic study on female of four species of the genus Coniopteryx (Neuroptera: Coniopterygidae) from China. Entomotaxonomia, 29: 19-25.
|
| ○ |
Ward, P. H., 1961. Parasemidalis annae (Enderlein, 1905) a synonym P. fuscipennis (Reuter, 1894) (Neuroptera: Coniopterygidae). Entomologist’s Gazette, 12: 114.
|
| ○ |
Ward, P. H., 1965. Some Ephemeroptera, Neuroptera and Trichoptera collected by mercury vapour light trap in a Hertfordshire
garden. Entomologist’s Gazette, 16: 169-174.
|
| ○ |
Ward, L. K., 1970. Aleuropteryx juniperi Ohm (Neur. Coniopterygidae) new to Britain feeding on Carulaspis juniperi Bouche (Hem., Diaspididae). The Entomologist’s Monthly Magazine, 106: 74-78.
|
| ○ |
Weidner, H., 1972. Die Entomologischen Sammlungen des Zoologischen Instituts und Zoologischen Museums der Universität Hamburg.
VIII Teil. Insecta V. Mitteilungen aus dem Hamburgischen Zoologischen Museum und Institut, 68: 107-134.
|
| ○ |
Wesmael, C., 1836a. Description d’un nouveau genre de Névroptères, famille des Planipennes, tribu des Hémérobins. Bulletins de l’Académie Royale des Sciences et Belles-Lettres de Bruxelles, 3: 166-168.
|
| ○ |
Wesmael, C., 1836b. Addition à la note de M. Wesmael, insérée dans le bulletin précédent. Bulletins de l’Académie Royale des Sciences et Belles-Lettres de Bruxelles, 3: 214-215.
|
| ○ |
Wesmael, C., 1841. Notice sur les Hémérobides de Belgique. Bulletins de l’Academie Royale des Sciences et Belles-Lettres de Bruxelles, 8(1): 203-221.
|
| ○ |
Westwood, J. O., 1834. On Coniortes, a new British genus of Neuroptera belonging to the Hemerobiidae. Transactions of the Royal Entomological Society of London (Proceedings), 1: xxvii. http://dx.doi.org/10.1111/j.1365-2311.1837.tb03167.x |
| ○ |
Westwood, J. O., 1840. Order V. Neuroptera Linn. In: Synopsis of the genera of British insects. London: 46-48.
|
| ○ |
Whalley, P. E. S., 1980. Neuroptera (Insecta) in amber from the Lower Cretaceous of Lebanon. Bulletin of the British Museum of Natural History, Geology, 33: 157-164.
|
| ○ |
Whiting, M. F., Carpenter, J. C., Wheeler, Q. U. & Wheeler, W. C., 1997. The Strepsiptera problem: Phylogeny of the holometabolous
insect orders inferred rom 18S and 28S ribosomal DNA sequences and morphology. Systematic Biology, 46: 1-68. http: //dx.doi.org/10.1093/sysbio/46.1.1
|
| ○ |
Williams, C. B., 1913. Coniopterygid cocoons. Transactions of the Royal Entomological Society of London (Proceedings), 61: lx. http://dx.doi.org/10.1111/j.1365-2311.1913.tb03108.x |
| ○ |
Willmann, R., 1990. The phylogenetic position of the Rhachiberothinae and the basal sister-group relationships within the
Mantispidae (Neuroptera). Systematic Entomology, 15(2): 253-265. http://dx.doi.org/10.1111/j.1365-3113.1990.tb00316.x |
| ○ |
Willmann, R., 1994a. Die phylogenetische Position ursprünglicher Mantispidae (Insecta, Planipennia) aus dem Mesozoikum und
Alt-Tertiär. Verhandlungen des Naturwissenschaftlichen Vereins in Hamburg (N.F.), 34: 177-203.
|
| ○ |
Willmann, R., 1994b. Raphidiodea aus dem Lias und die Phylogenie der Kamelhalsfliegen (Insecta: Holometabola). Paläontologische Zeitschrift, 68: 167-197. http://dx.doi.org/10.1007/BF02989439 |
| ○ |
Winterton, S. L., 2003. Molecular phylogeny of Neuropterida with emphasis on the lacewings (Neuroptera). Entomologische Abhandlungen, 61: 158-160.
|
| ○ |
Winterton, S. L., Hardy, N. B. & Wiegmann, B. M., 2010. On wings of lace: phylogeny and Bayesian divergence time estimates
of Neuropterida (Insecta) based on morphological and molecular data. Systematic Entomology, 35: 349-378. http://dx.doi.org/10.1111/j.1365-3113.2010.00521.x |
| ○ |
Wise, K. A. J., 1991. Distribution and zoogeography of New Zealand Megaloptera and Neuroptera. Records of the Auckland Institute and Museum, 28: 211-227.
|
| ○ |
Wise, K. A. J., 1992. Distribution and zoogeography of New Zealand Megaloptera and Neuroptera (Insecta). In: Canard, M., Aspöck,
H. & Mansell, M. W. (eds.). Current Research in Neuropterology (Proceedings of the Fourth International Symposium on Neuropterology, Bagnères-de-Luchon
1991). Toulouse: 393-395.
|
| ○ |
Withycombe, C. L., 1922a. Parasemidalis annae, Enderlein, a coniopterygid new to Britain, with notes on some other British Coniopterygidae. The Entomologist, 55: 169-172.
|
| ○ |
Withycombe, C. L., 1922b. The wing venation of the Coniopterygidae. The Entomologist, 55: 224-225.
|
| ○ |
Withycombe, C. L., 1923. Notes on the biology of some British Neuroptera. Transactions of the Royal Entomological Society of London, 70(3-4): 501-594. http://dx.doi.org/10.1111/j.1365-2311.1923.tb02844.x |
| ○ |
Withycombe, C. L., 1924a. On two new species of Coniopterygidae (Neuroptera) from Egypt. Bulletin de la Société Royale Entomologique d’Égypte, 7: 140-151.
|
| ○ |
Withycombe, C. L., 1924b. Note on the economic value of the Neuroptera, with special reference to the Coniopterygidae. Annals of Applied Biology, 11: 112-125. http://dx.doi.org/10.1111/j.1744-7348.1924.tb05696.x |
| ○ |
Withycombe, C. L., 1925a. Some aspects of the biology and morphology of the Neuroptera. With special reference to the immature
stages and their possible phylogenetic significance. Transactions of the Royal Entomological Society of London, 72(3-4): 303-411. http://dx.doi.org/10.1111/j.1365-2311.1925.tb03362.x |
| ○ |
Withycombe, C. L., 1925b. A contribution towards a monograph of the Indian Coniopterygidae (Neuroptera). Memoirs of the Department of Agriculture of India, Entomological Series, 9: 1-20.
|
| ○ |
Yang, C.-k., 1951. New records of neuropterous insects from China. Coniopterygidae Enderlein, 1905. Annales Entomologici Sinici, 1: 341-347.
|
| ○ |
Yang, C.-k., 1964. Notes on Coniopterygidae (Neuroptera). I. Genus Coniocompsa Enderlein 1905. Annales Entomologici Sinici, 13: 283-286.
|
| ○ |
Yang, C.-k., 1974. Notes on Coniopterygidae (Neuroptera). II. Genus Conwentzia Enderlein. Acta Entomologica Sinica, 17: 83-91.
|
| ○ |
Yang, C.-k. & Liu, Z.-q., 1993a. Neuroptera: Coniopterygidae. In: Huang, C.-m. (ed.). Longqi shan dong wu. Zhongguo lin ye chu ban she. Beijing: 225-226.
|
| ○ |
Yang, C.-k. & Liu, Z.-q., 1993b. The genus Cryptoscenea new to China, and a species new to science (Neuroptera: Coniopterygidae). Entomotaxonomia, 15: 249-251.
|
| ○ |
Yang, C.-k. & Liu, Z.-q., 1994. New species and new record of Coniopteryginae from Guangxi (Neuroptera: Coniopterygidae).
Journal of the Guangxi Academy of Sciences, 10: 75-85.
|
| ○ |
Yang, C.-k. & Liu, Z.-q., 1999. Coniopterygidae. In: Huang, B.-k. (ed). Fujian kun chong zhi. Vol. 3. Fujian ke xue ji shu chu ban she. Fuzhou: 86-94, 155-157.
|
| ○ |
Zakharenko, A. V., 1987. [A new species of the genus Coniopteryx (Neuroptera, Coniopterygidae) from the Far East.] Vestnik Zoologii, 1987(5): 76. (en ruso).
|
| ○ |
Zakharenko, A. V., 1988. [New and little-known dustywings (Neuroptera, Coniopterygidae) from Kazakhstan and Middle Asia.]
Zoologicheskii Zhurnal, 67: 1248-1250. (en ruso).
|
| ○ |
Zakharenko, A. V., 1989. [Species of Neuroptera of the families Coniopterygidae and Hemerobiidae new to the fauna of the USSR.]
Vestnik Zoologii, 1989(2): 87. (en ruso).
|
| ○ |
Zelený, J., 1961a. A contribution to the identification of the family Coniopterygidae (Neuroptera) in Bohemia. Casopis Ceskoslovenské Spolecnosti Entomologické, 58: 169-179.
|
| ○ |
Zelený, J., 1961b. Differentiation of Conwentzia psociformis Curt. from Conwentzia pineticola End. (Neuroptera). Casopis Ceskoslovenské Spolecnosti Entomologické, 58: 380-383.
|
| ○ |
Zelený, J., 1962. A contribution to the knowledge of the order Neuroptera in Czechoslovakia. Casopis Ceskoslovenské Spolecnosti Entomologické, 59: 59-67.
|
| ○ |
Zelený, J., 1964. Aleuropteryx umbrata n. sp. a new species of Neuroptera from Moldavian SSR (Neuroptera, Coniopterygidae). Casopis Ceskoslovenské Spolecnosti Entomologické, 61: 327-329.
|
| ○ |
Zelený, J., 1971. Neuroptera, Megaloptera und Mecoptera aus Bulgarien. Acta Faunistica Entomologica Musei Nationalis Pragae, 14: 153-163.
|
| ○ |
Zelený, J., 2005. Neuroptera (Lacewings)]. In: Farkač, J., Králd, D. & Škopik, M. (eds.). Red list of threatened species in the Czech Republic. Invertebrates. Agentura ochrany přírody a krajiny ČR. Praha: 163-165.
|
| ○ |
Zimmerman, E. C., 1957. Insects of Hawaii: a manual of the insects of the Hawaiian Islands, including an enumeration of the species and notes on their
origin, distribution, hosts, parasites, etc. Vol. 6. University of Hawaii Press. Honolulu. xi + 209 pp.
|
| ○ |
Zimmerman, E. E., 1940. Studies of Hawaiian Neuroptera. Proceedings of the Hawaiian Entomological Society, 10: 487-510.
|
| ○ |
Zimmermann, D., 2005a. Im Dschungel von Borsten, Kämmen & Wachsdrüsen REM-Untersuchungen an zwei Coniopterygiden (Neuroptera,
Neuropterida). DGaaE Nachrichten, 19: 145-146.
|
| ○ |
Zimmermann, D., 2005b. Rasterelektronenmikroskopische Untersuchungen an Imagines zweier Spezies der Familie Coniopterygidae (Neuroptera: Insecta). Master’s thesis. University of Vienna. Vienna. 77 pp.
|
| ○ |
Zimmermann, D., Klepal, W. & Aspöck, U., 2009. The first holistic SEM study of Coniopterygidae (Neuroptera)-structural evidence
and phylogenetic implications. European Journal of Entomology, 106: 651-662. http://dx.doi.org/10.14411/eje.2009.081 |
| ○ |
Zimmermann, D., Randolf, S., Metscher, B. D. & Aspöck, U., 2011a. Phylogenetische und funktionelle Bedeutung des Tentoriums
der Neuroptera (Insecta). Entomologica Austriaca, 18: 159-160.
|
| ○ |
Zimmermann, D., Randolf, S., Metscher, B. D. & Aspöck, U., 2011b. The function and phylogenetic implications of the tentorium
in adult Neuroptera (Insecta). Arthropod Structure & Development, 40: 571-582. http://dx.doi.org/10.1016/j.asd.2011.06.003 |
| ○ |
Zizzari, Z. V., Lupetti, P., Mencarelli, C. & Dallai, R., 2008. Sperm ultrastructure and spermiogenesis of Coniopterygidae
(Neuroptera, Insecta). Arthropod Structure & Development, 37: 410-417. http://dx.doi.org/10.1016/j.asd.2008.03.001 |
| ○ |
Zizzari, Z. V., Lupetti, P., Pantaleoni, R. A., Letardi, A. & Dallai, R., 2011. Sperm structure of some Neuroptera and phylogenetic
considerations. Italian Journal of Zoology, 78: 35-44. http://dx.doi.org/10.1080/11250003.2010.497503 |